ゲノムの発生
2016年8月1日
これから何回かにわけて、生物進化とその過程で生まれた情報について見ていく。この作業を通して生物進化を、様々な情報が生まれる過程として眺めてみたい。そしてこれらの情報を、「伝達性・コミュニケーション」と「環境の自己への同化」の観点から再検討することで、なぜ進化の究極に言語が誕生出来たのかに至る議論が出来ればと思っている。
前回述べたが、ある現象を特定のシグナルとして解釈できる生物機構がうまれると、この機構により解釈されることで、あらゆる現象は情報に変わる。この機構を理解することは当然生物進化の理解に欠かせないが、このシリーズでは、個体間、細胞間で伝達・コミュニケーション可能な情報に限って議論を進める。
元々、20世紀の情報理論は情報の伝達とコミュニケーションについての科学として生まれた。例えばシャノンの情報理論は、非物理的存在である情報が、伝達という観点から見ると、物理的物質(例えば電線)と量的な相互作用をすることができること、また情報が物理量による変化を受けてもコミュニケーションを成立させるための理論として始まった。また、選択された形質の子孫への「伝達性」はダーウィン進化論の中核をなす概念だ。生物にとっての情報は必ずしも個体間で伝達される必要はないが、今回は伝達・コミュニケーション可能な情報に限って議論する。
しかし、生物間で伝達とコミュニケーションに使える情報はゲノムだけではない。高等動物になると、ゲノム以外にも他の種の生物に伝えることのできる情報は数多く存在する。例えば言語も人間(生物)が持つ情報媒体だ。言語を獲得することで、私たちは自己を身体から解放し、ほぼ全ての人にほとんど無限の情報を伝えることができ、さらには他の個体と瞬時にコミュニケーションすることが可能になった。
このことを哲学者ヘーゲルはうまく表現しているので引用しよう。
「言葉は、自分を自分自身から分離する自己である。これは、純粋な自我=自我という形で、自らの対象となり、この対象態のうちにこの自己としての自らを支えるとともに、またそのまま他者と合流しており、他人の自己意識でもある。」(ヘーゲル 精神現象論:樫山欽四郎訳))
この様に伝達性・コミュニケーション性から見ると言語は情報進化の究極にあるが、言語に至るまで伝達可能な他の情報が進化の過程で現れた。これらの情報がどこに向かって、どのように伝えられるのかを見ていくのがこれからの作業だ。
情報の進化を理解するためのもう一つの観点に「環境の同化」を選んだのは、環境を同化しようとする指向性が進化の原動力と考えているからだ。環境による選択と言ってもいいのだが、ゲノム以外の情報が加わってくると、もっと積極的な環境の取り込みが起こっている。従って、ゲノム以降新しく生まれた様々な情報が環境を同化する過程を見ることで、進化の力学の本態を知ることができる。
環境の同化を考えることで、「進化は多様化した個体を環境が選択すると過程」といった紋切り型の議論を排して、進化の本当の力学を理解できるようになると期待している。そのため、個々の情報が進化にどう関わり得るかについても丁寧に検討してみたい。
伝達性を考えるとき、一つ注意してほしいのが、環境の変化を通して個体の活動が子孫に伝達できる点だ。あらゆる生物は様々な大きさの領域に拡がる環境を変化させる可能性を持っている。生物により変化させられた環境は、回り回って子孫の生存を制約できる。すなわち、生物による環境変化に関わった全ての情報は、こんどは環境からの制約という形で子孫に伝わる。少しわかりにくいと思うが、例えば言語情報を介する高次機能によりこれまでなかった新しい生活環境(例えば都市)が生まれたとしよう。こうして出来た都市は影響力のある環境として、子孫の生存や進化を制約する。これは人間だけの問題ではない。都市から排出されるスモッグで黒い羽を持つ蛾が都市では優勢になるという有名な「工業暗化」の話は、生物同士が環境を変化させることで、それぞれの進化に影響した例と言える。この生命の「歴史性」の問題は、進化を考える上でも重要な問題で、ゲノム以外の情報の進化への影響を扱う際常に留意する必要がある。
図1にこれから扱おうと思っている情報を図示したが、1)ゲノム、2)フェロモン、3)エピゲノム、4)神経回路と脳の高次機能、5)言語、そして最後に6)バーチャルリアリティーに限ろうと思っている。図に示すように、これらの情報を全て持つ生物は現在地球上には人間しか存在していない。また、地球上で生まれた伝達可能な情報は全て生命活動と連関しており、生命誕生と同時に生まれたゲノムから、約2万年前に誕生する言語まで、それぞれの情報の誕生は、生命の進化の方向性を大きく変化させ、また地球自体を作り変えてきた。
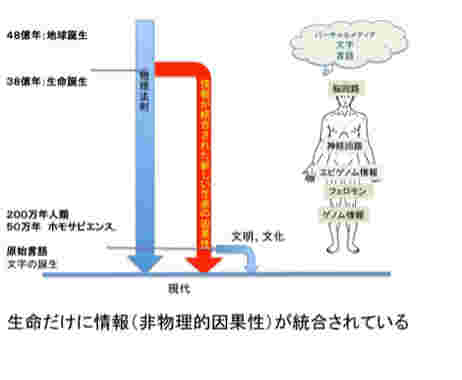
図1 進化から生まれた様々な情報:今回から順に扱おうとしている生命情報。これらは、38億年前のゲノム誕生と、約2万年前の言語誕生の間の生物進化過程で生まれた。
次回から図1に示した、生命情報、というより地球上に順番に現れた様々な情報進化として生物進化を眺め、それぞれの情報を伝達性と、環境同化性の観点から整理してみたいと思う。
ただ、地球に現れた最初の情報、ゲノムに関しては、これまで十分述べてきたので、今回手短に触れておくだけにする。
情報1:ゲノム
ゲノムを一言で定義せよと言われれば、私は「生殖を通して伝えることができる唯一の生物の情報」と定義している。生殖を通してだけ伝わるということは、ゲノムの伝達性は、ほぼ子孫に制限されていることを意味している。すなわち、複製を通して情報が子孫に共有され、それにより生きるためのメカニズムも共有される。
しかし他の生物の遺伝子が一部、あるいは丸ごと統合した名残を現在の生物にも見ることが出来る。例えばミトコンドリアやクロロフィル、あるいは水平伝搬でゲノムに入り込んだDNA断片だ。このため、生殖でしか伝えられないと言ってしまうのは言い過ぎとは思うが、高等動物になればなるほど「生殖を通してのみ伝えられる」という条件に従うようになる。
ただ忘れてならないのは、「生殖を通してのみ子孫に伝わる」というメカニズムと、他の個体の遺伝子を同化する過程を融合させた、「性生殖」という方法が生まれたことだ。
生物学的「性」とは、まさに個体間の遺伝子情報交換のことだ。これは原核生物にも存在し、一つの個体から他の個体に情報が移り、そこで相同組み替えが起こることで、異なる個体間の情報が一つの情報に作り直される。さらにこの方法でゲノム自体のコミュニケーション(相同組み換え)も可能になった。すなわちゲノムは伝達可能でコミュニケーション可能な情報になった。
このスッキリした定義は20世紀になり、少し危なっかしくなった。すなわち、人為的にゲノムを改変することが自由に行えるようになった結果、ゲノムの伝達性に関する制約条件は新たに変わろうとしているが、これについては言語を考えるときにもう一度議論したいと思う。
次に進化を環境の自己への取り込みという観点から見たとき、ゲノムが取りうる方法について見てみよう。
まず環境をRNAやDNAとして直接取り込むことが可能だ。この最たる例が、ミトコンドリアや葉緑体で、最初は体内の寄生生物として存在していたミトコンドリアの遺伝子が徐々に宿主のゲノムに移行して、宿主の自己に一体化していく過程は、まさに環境の取り込みとして見るとわかりやすい。
ミトコンドリアに限らず、環境 を情報として直接取り込む遺伝子伝搬は誕生初期の生物では普通に起こっていたと考えられる。ただ、この方法で直接環境からゲノムを取り込むことは、原始ゲノムから、単細胞、そして多細胞個体へと生物の自己の範囲が拡大するにつれてその頻度は下がっていった。特に多細胞動物になると、自己の制約から完全に独立した情報をそのまま取り込むことは、弊害の方が多くなったのだろう。この危険は、侵入した外来のゲノムの活性化を抑え込むエピジェネティックなメカニズムが進化しているのを見ると理解できる。
このことから、ゲノムは外来のDNA(情報)を容易に受け入れることができるが、自己という制約とは無関係に生まれた情報をそのまま取り込むことは、それがいかにゲノムの多様化に寄与するとしても、危険を伴うことがわかる。
例外として、既に述べたが性生殖という様式により、他のゲノムの断片を、別のゲノムに移すことが可能になっている。この場合、相同組み換えという機構が用いられるが、自己の制約の中で(相同性)ゲノム同士の情報が交換されるという様式が厳しく守られていると見ることが出来る。
もちろん、現在もなおトランスポゾンがゲノムの多様化に寄与する例はあるが、ゲノムが環境をとりこむための過程は、
- 1)変異と性を介したゲノムの多様化、
- 2)多様化した情報の翻訳、
- 3)翻訳された形質の選択
を繰り返すことで進む。
繰り返すが、この稿では選択を環境の同化として捉えていく。誤解を恐れず言ってしまうと、これはキリンの長い首を、丈の高い木の幹という環境が同化した結果と考えることだ。
同じような同化の例として亀の甲羅の進化についての最近の研究を紹介して終わろう。論文のタイトルは「Fossorial origin turtle shell (かめの甲羅は穴掘りが起源)」で、7月号のCurrent Biology (Vol 26:p1887, 2016)に掲載されている。
亀の甲羅の進化は、通常頭などの体の柔らかい部分を隠す硬いシェルターとして進化したと考えられてきた。例えば強い歯や爪を持った天敵の存在が、心臓や肺を守る硬い肋骨の進化を促したとでも考えればいいのだろうか。すなわち天敵のいるという環境圧が硬い甲羅を選択したと考えられていた。
ところがこの論文では、最初からシェルターを肋骨から作ろうとすると、運動能力や呼吸機能が低下し、逆効果になる。一方、亀の甲羅は最初穴掘りに適した骨格として進化したと考えると、2億年から2億5千万年前に生息していたと考えられるカメが、強い肋骨に支えられた前腕、長い爪など穴掘りに適した骨格を持っていることとも話は合うと主張している。
この主張の真偽はともかく、シェルターの必要性の観点から見ると、先に甲羅を発達させるより、確かに穴を掘った方が早い。そして、その後カメだけはこのシェルターとしての穴を、甲羅という形で自己に同化することに成功したと考えると、まさに環境が自己に同化している。
進化での選択を、環境の同化と置き換えてみるのも悪くないと思っていただいただろうか。
さて、次回はまずフェロモンから始める。
[ 西川 伸一 ]
情報の進化:1 フェロモンから見る情報の伝わり方と意味
2016年8月15日
情報を伝えるというと、私たちはすぐ何らかの媒体で表現された情報を個体間でやりとりすることを考えるが、原始ゲノムから細胞へと自己を拡大したばかりの単細胞生物では、情報の伝達とは、個体と一緒に同じ情報が増殖することで、この結果、同じ情報を共有した異なる個体が生まれる。もちろんゲノムの複製はゲノムの多様化につながり、同じ情報というのは正確ではないが、ゲノムの伝達のためには複製以外の道はない。
こう言い切ると、「原核生物には水平遺伝子伝播で文字どおりDNAを媒体とした情報を伝えるメカニズムがあるではないか」と指摘を受けそうだ。しかし、ウイルスやプラスミドのような寄生体のDNAを別にすると、水平伝播で伝えられるDNA断片を情報と呼ぶことには問題がある。というのも、多くの場合DNAがゲノムに取り込まれても、取り込んだ方の個体がその情報を解釈する仕組みを持っていないことが多い。例えて言えば、私にとってロシア語の文章が情報になりえないのと同じだ。逆に、解釈できない異なる種由来のDNAを闇雲に取り込んでしまうと、情報が伝わるどころか、自らのゲノムの完全性(genomic integrity)が損われるほうが問題になる。事実、原核生物のゲノムは、環状化し、相同性のないDNAが挿入されないようにして、外来のDNAの侵入に備え、ゲノムのインテグリティーを守っている。
おそらくLUCA (Last Universal Common Ancestor:全生物の共通祖先)が誕生した最初の頃は、ゲノムがコードする分子の特異性も高くなく、少々の外来DNAの挿入は許容され、外来のDNAを積極活用する余地があったかもしれない。しかし、時間をかけて最適な機能分子が進化し、自己の範囲の中で(実際には細胞の中で)厳密な分子ネットワークが生まれると、変異や外来DNAの侵入は、進化には都合が良くても、個体にとってのリスクは大きい。
ゲノムのインテグリティーを守るためのDNA修復機構は、地球上の全ての生物に備わっており、LUCA誕生後おそらく急速に発展したはずだ。自らのゲノムと相同性を持つ場合にのみDNAをゲノム内に受け入れる仕組みは、まずゲノム修復を転用するシステムとして生まれ、その後DNA情報を交換する細胞の「性」の基本メカニズムとして発展する。
いずれにせよ、性を通して自分と相同性が高いDNA情報だけを伝える仕組みが発達し、ゲノム間の部分交換が可能になるまでは、他の個体からDNAが情報として伝えられることはなかったと言っていい。
ゲノムの新しい世代への伝達を生殖(増殖)に限ることにより、一個の個体由来の子孫がほぼ同じ分子を共有できることは重要だ。すなわち、細胞内外の同じメカニズムを全て共有する、自分のコピーが数多く周りに存在することで、様々な刺激に集団が同じように反応するポテンシャルが用意される。すなわち、生命を成り立たせている制約自体を集団へと増殖させることが可能になっている。
フェロモン
このような準備の後、新しく生まれた情報伝達機構が、フェロモンとエピジェネティック機構と言える。フェロモンは現存の原核生物や古細菌から存在しており、一方完全なエピジェネティック機構は真核生物から見られることを考えると、おそらくフェロモンの方が早く誕生したのだろう。
原核生物のフェロモンが最もよく研究されているのがQuorum sensing(クオラムセンシング)と呼ばれる過程だ。そこで、先ずこのQuorum sensingが何かから見ていこう。
Quorum Sensing
Quorumは日本語で定足数と訳される。すなわち、何かを行う決断するために必要な個体数のことだ。図1に示すように、Quorum sensingは個体濃度を常に測定して、個体数が一定数を超えた場合のみ、特定の分子発現(図では赤丸)のスウィッチをオンにする仕組みだ。
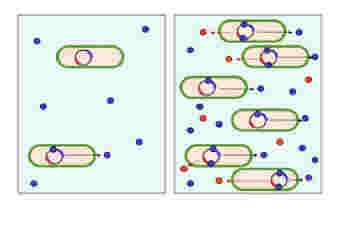
図1 Quorum Sensing 個体数を検出して(青丸の濃度)、一定の個体数に達したときだけ分子(赤丸)が発現する。このため定足数検出という名前がついた。(wikimedia
commonsより)
実はこの現象が最初に明らかになったのは、イカに住みついて光を発する発光バクテリアvibrio fisheriの研究からだ。すなわち、vibrio fisheriが蛍光物質を作るためには、菌がイカの体内で一定の濃度に達する必要があることがわかった。その後この仕組みが、様々な過程に使われていることが明らかになる。例えばQuorum sensingは細菌感染学でも重要な問題になっている。食中毒などのトキシンを分泌する事が出来る菌も、一定の菌数濃度に達しないと毒性を発揮しないことが知られている。このセンサーを破壊して、トキシンの分泌を止めることができると、有効な治療法になる。
Quorum sensingの分子メカニズムはほぼ完全に解明されている。このメカニズムには、定足数(菌濃度)を教えるための指標(インデックス)となる分子、その分子が一定濃度に達したことを感知する分子、そしてこの認識を様々な物質の生産につなげるための転写活性化メカニズムが必要になる。この分子機構については多くのバクテリアで解明が進んでいるので、ここではグラム陽性菌を例にQuorum sensingに関わる分子機構を示す。
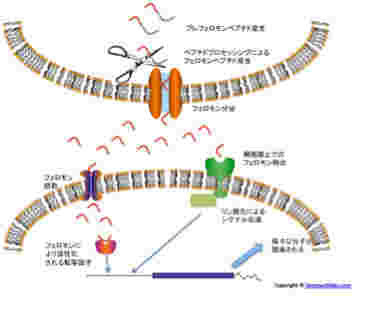
図2:グラム陽性菌でのフェロモンによるシグナル伝達経路。説明は本文参照。
この系で定足数の指標になる分子がフェロモンだ。グラム陽性菌では私たちのペプチドホルモンと一緒で、先ず長めのペプチドが作られ、それが細胞外へ分泌されるとき短く剪定される。この仕組みにより、ペプチドを作った同じ細胞にセンサーが存在していても、一度細胞外へ出たペプチドだけが刺激できるようになっている。
一方、グラム陰性菌ではペプチドの代わりにラクトンを核にした有機化合物を使っている。これはエストロジェン(Estrogen:女性ホルモン)などの核内受容体に対するリガンドと似ていると言えるかもしれない。
Quorum sensingが働く菌は常にフェロモンを作り続けているが、菌濃度が低いと細胞外のフェロモン濃度は検出感度以下でとどまり、フェロモン受容体は活性化されない。ところが菌数が増えてくると、それに応じて細胞外のフェロモン濃度が上昇し、受容体は活性化され、標的遺伝子の転写のスウィッチが入る。これがQuorum sensingのメカニズムだ。
どちらのタイプのフェロモンにも2種類のセンサーが特定されている。一つは、細胞内に入ってきたフェロモンと結合して、転写活性のスウィッチが入る転写因子で、もう一つは細胞表面上でフェロモンと結合し、リン酸化反応を介してシグナルを伝える細胞表面受容体タイプだ。この構造を見ていると、人間がホルモンとして使っている、核内受容体(例えばエストロジェン受容体)と細胞外受容体(例えば成長ホルモン受容体)などと同じタイプのメカニズムがバクテリアから存在していることがわかる。
メカニズムはこのぐらいにして、フェロモンシステムの進化を考えてみよう。まず、原核生物で見られるフェロモンシグナル伝達システムの基本構造は、その後の生物で普遍的に見られることから、フェロモン自体の進化については割愛して、LUCAがフェロモンシステムを進化させてきた過程について考えてみよう。
理解を容易にするため、ここではフェロモンが細胞内で転写因子に結合して転写を活性化させる系だけを考える。ここで使われている分子メカニズムは、まさに細胞内で合成された分子を使ってフィードフォワードループを動かす系と同じで、あらゆる生物で普遍的に見られる。ジャコブとモノーが発見したトリプトファンに結合してトリプトファンオペロンを抑制するトリプトファンレプレッサーは最も有名な例だ。ただ細胞内部にメッセンジャーがとどまるフィードフォワード回路とは異なり、そのまま、或いはプロセッシングを受けたあとフェロモンとして細胞外に分泌されることで、自分だけでなく他の個体にも作用できるようになっている。
メッセンジャーを細胞内だけでなく、細胞外に分泌できるようにして他の個体にも情報を伝達する進化により、それまで全く単独で生きてきた生物は自己の範囲を広げ、他の個体と情報を共有し、同じ行動を取る一種の社会行動が可能になった。

図3 バイオフィルム(wikimedia commonsより)。バクテリアが一定濃度に達してEPSを分泌しフィルムを形成、それをバクテリアが一種の住処として利用している。
Quorum sensingの意義を考えるとき、バクテリアがEPSと呼ばれるポリマーを分泌して集合体を作るバイオフィルム形成過程で、同じ機構が働いていることは興味深い。バイオフィルムは単独では決して形成できない。形成に必要な細胞数に達しているかどうかの情報共有をQuorum sensingは可能にしている。すなわち、自己の範囲を拡大する進化をここに見ることができる。さらに、バイオフィルムは、環境の自己への同化が起こっている例として見ることができないだろうか?
ではこのシステムで生まれた情報の特徴について最後に見ておこう。まず情報の媒体にコード性はない。しかし、記号論的にいうとフェロモンは個体数のインデックスになっている。そしてこのインデックスを解釈する仕組みを共有することで、インデックス自体とは物理的に関係のない物質を作ることが可能になっている。このように、フェロモンシステムはゲノムと異なりコード性は全くないが、代わりにインデックス記号性を持ち、これを介して同時代の他の個体と情報の共有とコミュニケーションが可能になっている。
バイオフィルム形成と同じ情報共有システムはその後、あらゆる単細胞動物で発達を遂げ、多細胞動物への道を開く一つの条件となり、生物の新しい情報の重要な類型となった。
次回は、単細胞生物から多細胞生物への進化に必要だったもう一つの情報伝達・コミュニケーションシステム、エピジェネティック機構について考える。
[ 西川 伸一 ]
情報の進化II クロマチン構造を媒体とした情報
2016年9月1日
フェロモンの次に議論する情報は「クロマチン構造を媒体とした情報」だ。この情報は一般的にエピジェネティックスと呼ばれることが多い。ただ、エピジェネティックスという言葉の指す範囲は曖昧で広い。このため、ここではエピジェネティックスを「クロマチン構造を媒体とした情報」に限定する。
まず簡単に「クロマチン構造を媒体とした情報」、エピジェネティックスについて簡単に解説する。
私たち人間は40兆個近くの細胞を持っているが、全て一個の受精卵が発生過程で、発生プログラムに従って様々な細胞に分化した結果だ。リンパ球のような一部の例外を除くと、何百、何千種類もの異なる細胞ができるためにゲノムの変化は全く必要ない。細胞の形は異なり、作られる分子は全く違っていても、私たちの体の中の細胞は全て同じゲノムを持っている。このゲノムの変化なしに異なる性質の細胞が発生し維持されるメカニズムがエピジェネティックスだ(図1)。
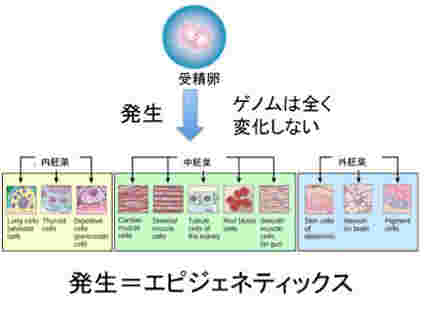
図1 発生ではゲノムは変化せず、クロマチン構造の変化だけが起こる。従って、ほとんどの発生過程は、エピジェネティックな過程と見ることができる。(図はWikiを拝借)
次にこの発生の結果何が可能になったか考えてみよう。何百種類の細胞がただ発生したというわけではない。例えば皮膚の細胞は皮膚細胞を作り続け、血液細胞は血液を安定に作り続けることが可能になっている。同じゲノムを持っている細胞同士なのに、決して皮膚の細胞が血液に変わることはない。もしそんなことが一定の確率で起こったら私たちの体の統合性はたちどころに破綻する。
これを支えるメカニズムがクロマチン構造を媒体とした情報=エピジェネティックスで、原則として真核生物で初めて見ることができる。このゲノムとヒストンと呼ばれるタンパク質が互いに協調することで形成されるのが、クロマチン構造で、それぞれの細胞種でパターンが異なる。また、細胞分裂でヒストンが外れても、もう一度同じように再構成できることから、クロマチン構造を繰り返し形成し直すための一種の鋳型ができていると考えると判りやすい(図2)。
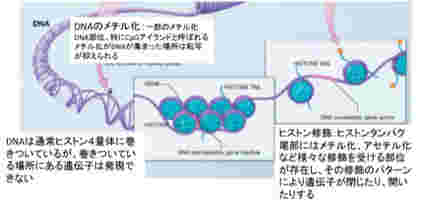
図2:クロマチン構造と修飾:ヒストンの少ないところはクロマチンが開いていると表現し、一般的にそこにある遺伝子は転写因子と相互作用する。一方、多くのヒストンが集まった部位はクロマチンが閉じていると表現し、ここにある遺伝子は転写因子から隔離されており、発現できない。(図はWiki
を拝借)
このクロマチン鋳型は、全ゲノムにわたって活性化部位と、不活性化部位を区別する印をつけることにより形成される。これらの印は、ノートの開きたい場所と、閉じておきたい場所につける付箋のようなものと考えるといい。
体の全ての細胞には、それが属する種類、すなわち皮膚細胞、血液細胞といった種類に対応する異なる鋳型が存在している。そして何よりも重要なのは、細胞分裂でこの鋳型も複製され、二つの娘細胞に伝わる。このような性質から考えれば、エピジェネティックスをクロマチン構造を媒体とした情報と呼ぶことは何の問題もない。また、この鋳型があるおかげで皮膚細胞は間違って血液細胞や脳細胞になる心配なく、皮膚細胞を作り続けることができる。実際、この鋳型を維持するメカニズムが狂うと、皮膚細胞が思いもかけない細胞へ分化するといった乱れが生ずる。ガンで見られる異型細胞の出現はこの例だ。
ではこのクロマチン鋳型とは何で、情報としてどのような役割を担っているのか見てみよう。すでに図2に図示したように、ゲノムは細胞の種類に応じてヒストンが詰まった閉じた状態と、結合しているヒストンが少ない開いた状態にわかれている。
この開いた状態、閉じた状態を決めているのが、染色体と結合しているヒストンのN末端にある領域に存在する数種類のリジン残基のメチル化とアセチル化、そしてDNA自体のメチル化だ。このゲノムの場所特異的なヒストン修飾パターンやメチル化DNAパターンが鋳型として子孫に伝わり、繰り返し働く。これを可能にするのが、発生過程で細胞分化に合わせて鋳型を書き込むメカニズムと、細胞分化のあと完成した鋳型を維持するメカニズムだ。
このメカニズムについての理解は最近急速に深まってきたとはいえ、今も研究途上の分野なので、ここでは解説は省く。ただ、多能性幹細胞を始め、様々な培養細胞のクロマチン鋳型を精密に調べる研究から、
- 1)細胞が分裂を繰り返しても同じ鋳型が子孫細胞に受け継がれること、
- 2)多能性幹細胞から様々な細胞系列が分化する過程で、各細胞に特異的なクロマチン鋳型が形成されること、
- 3)分化細胞をそのまま培養しても普通はクロマチン鋳型が変化することはないが、iPS誘導のように特殊な条件下ではリプログラムが可能で、この時実際に変化するのはこのクロマチン鋳型であること。
などが明らかになった。
この結果は、発生や成長で細胞分化のシグナルに応じてクロマチン鋳型を書き換えるメカニズムと、書き換えたクロマチン鋳型を安定に維持するメカニズムが間違いなく存在することを示している。すなわち、塩基配列のようなコード性は存在しないが、クロマチンのパターンを解釈して複製できることから、エピジェネティックスをフェロモンの次に誕生した情報と呼ぶことができる。
最後にクロマチン構造を媒体とした情報の特徴を、ゲノム、フェロモンと比較しながら見てみよう。
前回見たように、フェロモンはリアルタイムの個体間のコミュニケーションを可能にした全く新しい情報で、ゲノムだけがどれほど複雑化してもなしえない可能性を拓いた。前回このフェロモンの役割について、単位あたりの個体数を感知して一定の反応を起こすquorum sensingを例に説明した。ただフェロモンが活躍するのはquorum sensingだけではない。研究が進むもう一つの領域が、バクテリア同士がプラスミドを受け渡すときに働くフェロモンだ。これによって薬剤耐性が多くのバクテリアに伝搬するため、医学にとっては重要な分野になっている。
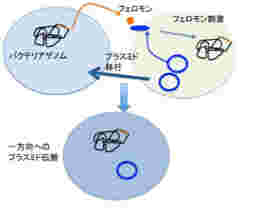
図3:フェロモン刺激によりプラスミドを持つバクテリアから持たないバクテリアへのプラスミドの伝搬が起こる。このメカニズムで薬剤耐性遺伝子が伝播される例が多く知られており、医学でも問題になっている。
図3に示す腸球菌の系では、フェロモンはバクテリアのゲノムにコードされているが、その受容体は薬剤耐性を持つプラスミドにコードされている。この結果、フェロモンに反応するプラスミド(+)バクテリアと、フェロモンを分泌するプラスミド(ー)バクテリア同士が特異的な接合を起こし、プラスミド陰性のバクテリアにプラスミドを渡すことが可能になっている。実際には、プラスミド(+)バクテリアが自分のフェロモンに反応しないよう、うまく抑制システムができているが、詳細は省く。
この現象からわかるのは、フェロモンが引き金となって、DNAを媒体とする情報のやりとりが行われ、この結果、効果が一過性のフェロモンのシグナルが、遺伝子変化として記録されることだ。すなわち、一過性のフェロモンの効果を長期間保持するためには、バクテリアでは遺伝子の変化として記録する必要がある。しかし、プラスミドのような限られた情報の伝搬は簡単でも、ゲノムを書き換えたりすることは難しく、出芽酵母のような一部の例外を除いて、同じ書き換えを繰り返すことは難しい。
一方真核生物への進化により、一過性に誘導される遺伝子発現状態をもとに、クロマチン鋳型を形成する能力、すなわちエピジェネティック過程が誕生すると、遺伝子の書き換えなしに、外界からのシグナルを特定のクロマチン構造として書き残すことが可能になる。
これをバクテリアがゲノムに記録する場合と比べると、外来の情報を持続的に記録するという点では同じだが、
- 1)記録が書き換え可能
- 2)多くのシグナルに対応して記録が可能。
- 3)同時に多くの個体がシグナルを共有し、それを同じ記録として維持することが可能。
などの、ゲノムでは不可能とは言えなくとも、簡単ではない記録が可能になっている。
これがクロマチン構造を媒体とする情報の誕生により生まれた可能性で、この獲得には20億年、生物の進化全過程の半分以上の時間が費やされている。次回は、この新しい情報の進化と、この情報がもたらしたインパクトについて考える。
[ 西川 伸一 ]
外界の情報を記憶するためのエピジェネティックス
2016年9月27日
前回「クロマチン構造変化を媒介とする情報(<エピジェネティックス)」が、
1)情報の書き換えができる、2)様々な外界のシグナルの影響を比較的迅速に記憶できる、3)異なる個体間で同じ情報を共有して同じ反応を可能にする、の3点で、ゲノムを媒体とする情報と異なることについて述べた。今回はこの違いについてもうすこし具体的に掘り下げる。
これを読んでいる読者のほとんどは、ルイセンコの名前を聞いても誰かわからないだろう。ルイセンコは、メンデル遺伝理論だけでは説明がつかなかった植物の現象を捉えて、獲得形質は遺伝することを主張したソビエトの農学者だ。彼を政治的に支持したスターリン政府の後ろ盾を得て、論敵を逮捕・追放したことで、科学的議論に政治を介入させたとして科学史に大きな汚点を残した人物だ。彼の理論は我が国にも様々な影響を残したようだが、これは私が大学に入学するより前の話だ。幸い、ルイセンコと彼の理論をめぐる我が国での議論については本館名誉顧問の岡田節人先生が生命誌ジャーナルに紹介されているのでぜひ読んでほしい。
さてルイセンコがメンデル遺伝学の反証の根拠にしたのが「春化(春処理)」と言われる現象で、植物を低い温度に晒すことで開花時期が調節できるという現象だ。低温という環境で獲得した形質が、世代を超えて伝わる場合もあることから、獲得形質が遺伝すると考えたようだ。結局エピジェネティックス(遺伝した後)の問題で、ゲノム自体の変異を基盤とする遺伝学の問題でないことが明らかになる。このように、春化メカニズムは、エピジェネティックス、すなわちクロマチンの構造を介した情報の問題として現在解明が進んでいる。外界からのシグナルがクロマチン構造を媒体とする情報として書き換えられるメカニズムを理解するのにうってつけの例なので、まずこの春化とそのメカニズムについて見てみよう。
各々の植物は発生過程で開花を調節する機構を確立する。これには温度や日照時間を感知しながら様々な遺伝子の発現を調節する複雑なメカニズムが関与する。話をわかりやすくするために、実験のためのモデル植物として研究されているシロイヌナズナの開花を抑制するマスター遺伝子FLCに絞って春化の現象を解説する。
まず越冬一年草型シロイヌナズナのライフサイクルを見てみよう。秋に発芽、ロゼット型葉を形成してゆっくり成長しながら越冬する。その後、茎が伸びて開花、そして夏になると種ができる。この間の開花抑制遺伝子FLCの発現を見てみると、発芽時の秋には最も高いレベルにある。しかし、ロゼットが形成される冬には急速にそのレベルが低下し、春の開花期には0になる。開花が終わると夏に向けて急速に発現レベルが回復する。
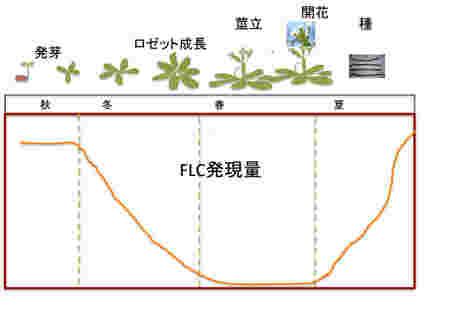
図1 シロイヌナズナの開花と、開花抑制遺伝子FLCの発現量。説明は本文参照。花と種の写真はWikipediaより。
FLCが開花抑制に関わることを頭において発現の季節パターンを見ると、開花の起こる春以外は開花抑制遺伝子であるFLCを発現させ、間違った時期に開花が進まないようできているのがわかる。すなわち、秋から冬にかけて気温が下がり始まるのを合図に、FLCの発現を安定に抑制するための準備を始め、この状態で急速に温度が上がると、開花を優先した植物の発生が進み、開花する。その後、種ができ、発芽するまでにFLCの発現を戻して開花を抑え、植物はエネルギーを成長に使うことが可能になっている。
次に、このパターンの背景にあるメカニズムを細かくみていこう。この季節に合わせた発現の調節にはFLC遺伝子領域とそこに結合しているヒストンのメチル化パターンが大きな役割を演じている。すなわちFLCの発現を抑えるときはこの領域に巻きついているヒストン3のN末27番目のリジンがメチル化されており(H3K27me3)、一方発現が高いときは4番目のリジンがメチル化されている(H3K4me3)(図2)。すなわち、クロマチンが閉じて転写が抑制されるか、あるいはクロマチンが開いて転写が行えるかは、ヒストンの標識の違いによって決まる。
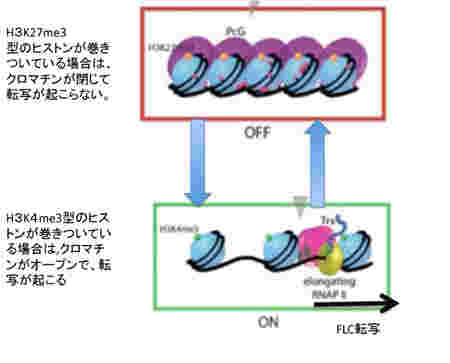
図2 ヒストンのメチル化によるクロマチンの構造のスウィッチ。図はES細胞分化時に起こるクロマチン構造変化についての説明をStem Bookサイトより転載。(http://www.stembook.org/node/585.html)
つまり、FLCに巻きついているヒストンのメチル化の異なる2つの状態を温度スウィッチでシフトできるようになっている。このようにクロマチン構造を媒体とする情報の誕生は、外界の変化を細胞自体の記憶に変えることを可能にした。
このヒストン標識によるクロマチン構造のスウィッチのメカニズムについてはずいぶんわかってきたが、複雑でその全像について解説するには本1冊必要になる。とりあえず、図にあるように、クロマチンの構造を維持する鍵になる分子が、ポリコム複合体とTrithorax複合体であることを理解しておけば十分だろう。
H3K4のメチル化と、H3K27のメチル化パターンを転換させクロマチン構造を変えることで大きな領域の遺伝子発現を調節するメカニズムは、実は酵母から人間まで保存されているメカニズムだ。事実、ポリコム複合体やTrithorax複合体という名前はショウジョウバエ突然変異の研究が由来となっている。例えば、ショウジョウバエから哺乳動物まで、体の前後軸に合わせて体節の特異性を決めているHox遺伝子が集まる大きなゲノム領域のどこまでを活性化し、どこまでを不活化するかは、まさにこのクロマチン構造変化により調節維持されている。
図3は哺乳類の発生でのHox遺伝子領域のヒストン標識パターンを図示しているが、Hox遺伝子の発現はクロマチン構造の変化で調節され、植物とほぼ同じメカニズムが使われていることがわかる(図3)
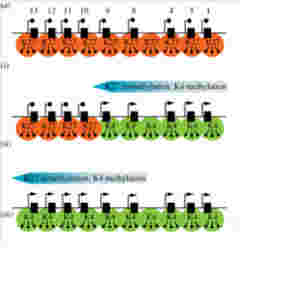
図3 クロマチン構造変化によるマウスHoxa1-13遺伝子の発現調節。内部細胞塊のような多能性幹細胞ではHox領域は完全にH3K27me3ヒストンで標識され、発現が抑えられている。前から後ろの体節に移行するのに合わせ(i)(ⅱ)(ⅲ)、Hoxa1からH3K4me3ヒストンは拡大していく。
出典(http://rstb.royalsocietypublishing.org/content/368/1620/20120367)
最後に、春化時のクロマチン構造の変化の際、他生物で見られない特徴について見ていこう。実は、FLC遺伝子発現を抑制するH3K27me3標識の範囲の量を調節できるようにできている。すなわち寒い時期が長いと、H3K27me3が結合するFLC遺伝子領域が拡大し、暖かくなってもこの抑制標識を外しにくくなる。これは、低温に長くさらされた植物の場合、開花が早くなり、開花期間は長くなり、さらに開花量も上昇するという量的な変化が、FLC遺伝子に結合するH3K27me3の量的な変化と相関していることを意味している。このように、外界からの情報を細胞内に量的な記憶として残すことができるのも、クロマチン構造を媒体とする情報の特徴だ。これをゲノム自体の調節機構で実現することはほぼ不可能だ。
春化を例に、クロマチン構造を媒体とする情報が、外界の情報を、細胞自体の情報に書き換えて記憶することを可能にしていることを示した。重要なことは、この記憶が書き換え可能で、量的にも変化させることが可能なことだ。進化が環境の自己への取り込みであるという観点から見ると、クロマチン構造を媒体とする情報は、ゲノムよりはるかに迅速に環境の自己への取り込みを可能にした新たな情報記憶の登場といえる。
今回例として使った春化では、環境の温度がクロマチン構造の情報として書き換えられる。一方、クロマチン構造の書き換えはフェロモンによっても誘導することができ、生命進化で誕生した新しい2種類の情報がここで出会う。すでに述べたように、フェロモンは個体間、細胞間のコミュニケーションに関わる情報と言える。一方、今回見てきたようにクロマチン構造を媒体とする情報は記憶媒体と言える。このように、コミュニケーションと記憶が揃った時、生命進化が大きく進む。こうして可能になったのが多細胞動物だ。次回はこの進化過程について考えてみる。
[ 西川 伸一 ]
コミュニケーション・伝達・記録
2016年10月3日
前回まで、ゲノム、フェロモン、クロマチン構造が媒介する情報と進化で生まれた新しい情報について見てきた。これらの情報を、伝達、コミュニケーション、記録という3つの情報機能について見直してみよう。
伝達、コミュニケーション、そして記録は情報が可能にする重要な機能だ。ゲノムはこれらすべての機能を併せ持っている。新しく生まれた情報には伝達機能は存在するが、フェロモンには記録という機能はなく、クロマチン構造を媒介とする情報はコミュニケーション機能を持たない。前回述べたが、フェロモンとクロマチン構造はゲノムの持つ情報を補完するように別々に誕生し、真核生物になると統合された情報システムへと発展する。そしてこの統合から生まれた新しい可能性が、多細胞動物の誕生の基盤になる。今回は、情報の記憶とコミュニケーション機能について整理をした上で、多細胞体制が生まれるまでに、生物の情報システムがどう発展したのか整理しよう。
まず、フェロモンやクロマチン情報が必要とされた背景、情報としてのゲノムの限界から見てみよう。
いうまでもなく、細胞が複製できるのは、ゲノムがDNAを記憶媒体として使って生物のオペレーションを指示する情報として働いているからだ。この記録機能と記録の伝達の原理については、DNAが2重螺旋構造をしていることが発見された時、解明されたと言っていい(図1)。
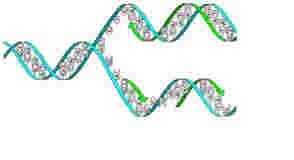
図1 2重螺旋の複製。2重螺旋が一本鎖にほどけ、それが2本の2重螺旋になることがDNAの構造から明らかになり、伝達と記録機能の原理が理解された。(wikipediaより)
ではコミュニケーション機能はどうだろう。複製されたゲノムは2つの娘細胞に伝わる。これは生殖を通しての情報伝搬だが、コミュニケーションではない。
ゲノム同士のコミュニケーションは基本的に相同組み換え機構を介して行われる。私たちの体細胞には、母親からの染色体と、父親からの染色体が存在するが、条件が整うと両方の染色体同士で相同組み換え(図2)が起こり、染色体の部分交換がおこる。これにより異なるゲノム同士のコミュニケーションが可能になる。
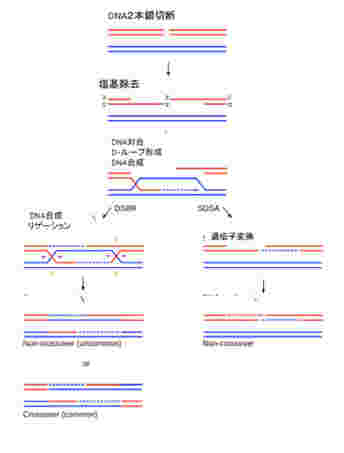
図2:相同組み換えを通した2つのゲノムのコミュニケーション(wikipediaより)
この機構はほぼすべての生物で保存されており、大腸菌から人間まで、異なる個体のゲノム同士のコミュニケーションにはこの機構が使われる。ただ、相同組み換えが起こるためには、DNAの切断が必要で、実際には放射線などによるDNA切断に対する修復機構として進化してきたと考えられる。通常の状態で相同組み換えが起こるためには、DNAを自然に切断する過程が必要で、細胞にとっては危険極まりない。このため多くの生物では、この機構を通したゲノム間のコミュニケーションが高い確率で起こる状況は限られている。例えば、人間では生殖細胞の減数分裂過程に限られ、体細胞で起こる頻度は低い。
このように、ゲノムの記憶、伝達、コミュニケーションの様式を見てみると、全てが生殖サイクルと完全に一体化していることがわかる。単細胞動物では、異なる個体同士が接合するとゲノムレベルのコミュニケーションが起こるのを観察できるが、コミュニケーションの範囲は1対1に限られており、一つのゲノム情報が同時に異なるゲノム情報とコミュニケーションすることはない(図3)
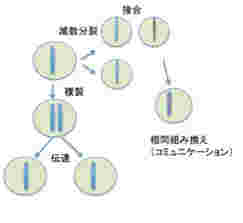
図3: ゲノムには情報が記録されており、複製を通して娘細胞に伝達される。また、接合、ゲノム間の相同組み換えにより、他の個体のゲノムとコミュニケーションできる。ただ、図からわかるようにゲノム情報の伝搬、コミュニケーションは全て一対一で個体(細胞)が対応しており、同時に複数の個体とコミュニケーションしたり伝達することはない。
ではなぜコミュニケーションが必要か、その利点を考えてみよう。ダーウィン進化を現代的に翻訳すると、様々な様式のゲノムレベルの多様化がまず起こり、ゲノムの多様化が形質の多様化として反影されると、この形質の環境適合性を指標に生殖優位性での差異が生じ、環境に適応した個体の比率が高まることで進化が進むと言えるだろう。このゲノムの多様化の一番の駆動力は、DNA複製に必ず伴うエラーだが、もう一段質の高い多様化の駆動力が相同組み換えによる多様化だ。
全くランダムな多様化と比べると、相同組み換えによる多様化は、ゲノムレベルのコミュニケーションを基盤にしている点が大きく異なる。通常これが起こるのは性生殖だが、この場合異なる環境で生きてきた個体がそれぞれ持っているゲノム同士の部分を交換し合う。視点を変えてこの過程を見てみると、その種が分離してから世代を重ねてきた異なる個体のゲノム同士、いわば環境によりすでに「テスト済み」のゲノム同士で情報交換が行われていることを意味する。それぞれの個体のゲノムがその個体が出会ってきた環境を同化していると考えると、この環境との相互作用の歴史が一つのゲノムの中へ再構成されているとみることができる。言葉を変えると、ゲノム間のコミュニケーションにより、より多くの環境との出会いが、ゲノムに同化される。これがゲノム間でコミュニケーションが行われることの重要な役割だ。
この様にゲノムには、情報としての全てが備わっている様に思える(事実それが進化の原動力となってきた)が、特定の生物集団の生存から見た時、ゲノムの対応能力には限界がある。すなわち、ゲノムだけで環境の変化に対応しようとすると、伝達やコミュニケーションが1:1に限られ、空間的にも、時間的にも急激な環境変化には対応できない。この限界を埋めるのが、フェロモンと、クロマチン情報だ。
詳しく検証したわけではないが、フェロモンは、細胞内の分子間相互作用ネットワークが、細胞外へ拡大したものと考えられる。フェロモンは接合の相手を惹きつけ、接合時の方向性(オスメスと考えて貰えばいい)を決める過程にしばしば登場する。これは1:1の個体間相互作用と言えるが、他にもクオラムセンシング(前々回)のように1:1の限界を超え、同時に多くの個体とコミュニケーションをとりあう手段を提供し、全ての個体が環境に対して同じように反応するための情報としても働いている。
この様に、生存環境に関する情報を他の個体と同時的に共有することを可能にしたのがフェロモンの機能だ。フェロモン自体の情報には記録機能がないため、フェロモンのシグナルは一時的な反応を誘導して終わるか、あるいはフェロモンの情報を引き金に、遺伝情報を書き換え(酵母の交配型の変換)や、プラスミドを介したゲノム情報のやりとり(前々回)を行うことで、経験が記録される。このゲノム自体の書き換えの代わりに、クロマチン情報の書き換え機構とフェロモンシグナルをリンクさせることもできる。実際、クロマチン情報の書き換えが、反応の多様性、伝達範囲の大きさ、伝達と記録の迅速性の点で、ゲノムの書き換えをはるかに凌駕しているため、これがその後の生物の主要な伝達様式になっていく。
前回みた様に、クロマチン構造を媒介とする情報は、書き換え、すなわち繰り返して記録が可能だ。このメカニズムが完成するのは、おそらく真核生物以降だが、これに必要な個々のメカニズムは原核生物や古細菌で進化してきた。事実DNAメチル化は原核生物から存在するし、ヒストンと相同性を持つタンパク質も古細菌から認められる。更には、遺伝子の相同性は欠如しているが、DNAと結合してヌクレオソーム構造を形成するためのタンパク質は原核生物にも存在している。
この様なヌクレオソーム構造が広く認められることは、プラスミドの様な小さなゲノムは別として、メガベース以上の大きさのゲノムは裸のままで存在することが不可能であることを示している。実際、バクテリアやアルケアでは、1mmの長さのDNAを1ミクロンの大きさの細胞内に安全に収納している。その上に、収納した全ゲノムをなんども複製することが求められる。ヒストンの様な、DNAをコンパクトに巻き取る仕組みなしには、ゲノムサイズの急速な増大はありえなかっただろう。
真核生物の誕生にはヒストン修飾を基盤とするクロマチン構造制御機構、分裂時の染色体分離に必要なチュブリン、そして形態や細胞分裂を制御できる細胞骨格分子アクチンが必要だが、この3種類のタンパクは同じ分子からできてきたと考える魅力的な仮説がある。すなわち、いずれもリン酸化やアセチル化による修飾を介した細胞の構造変化に関わる。この考えを採用した、真核細胞進化過程について図3にまとめてみた。
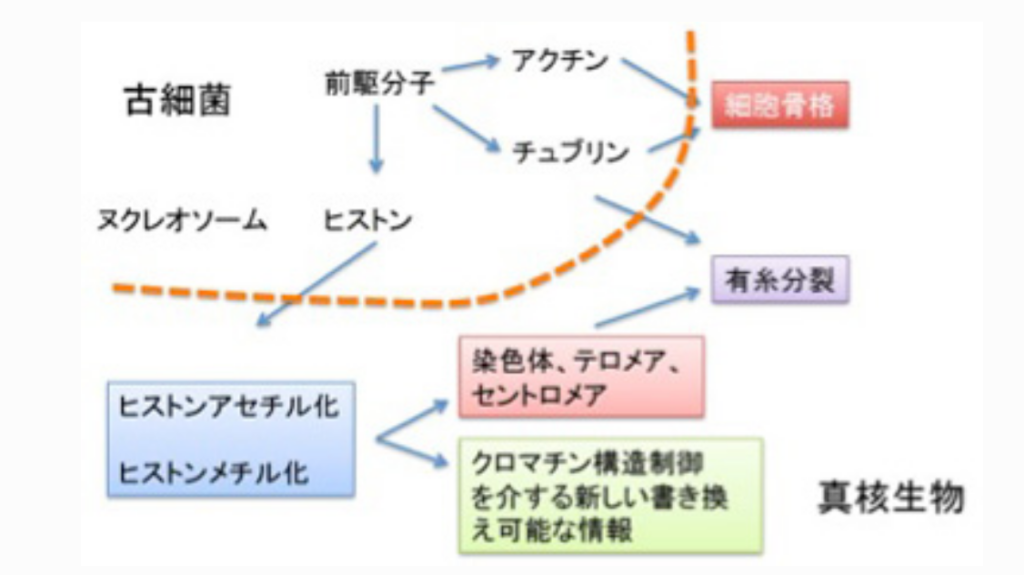
図3:古細菌から真核生物への進化をヒストンから眺めてみる。
ここでは話をヒストンに限るが、最初DNAをコンパクトに折りたたみ保護するヌクレオソーム形成分子として誕生したヒストンは、N末端に集まるリジン残基のアセチル化/脱アセチル化、メチル化/脱メチル化を介する修飾により、単純なヌクレオソームから、各ゲノム領域を構造的に標識するクロマチン構造へと発展した。これにより、テロメアやセントロメアのように、有糸分裂に必要な条件が整った染色体が生まれる。そしてゲノム領域を構造的に標識するヒストンの同じ能力が、遺伝子発現調節に使われるようになったのが、クロマチン情報だ。
春化の例でクロマチン情報は、環境のストレスを、多くの個体が同時に、同じシグナルとして感知し、このシグナルを同じ様に記録することを可能にしている。このようにクロマチン情報は複数の個体が同時に共有できる、ゲノムではなしえなかった情報と言える。
最初環境ストレスに対する反応として誕生したクロマチン情報の書き換え機構に、フェロモンによる細胞間情報共有機構が統合されると(図4)、それ自身では互いに未完成であった情報が補い合って、コミュニケーション、伝達、記録のすべて備わった新しい情報システムが完成する。
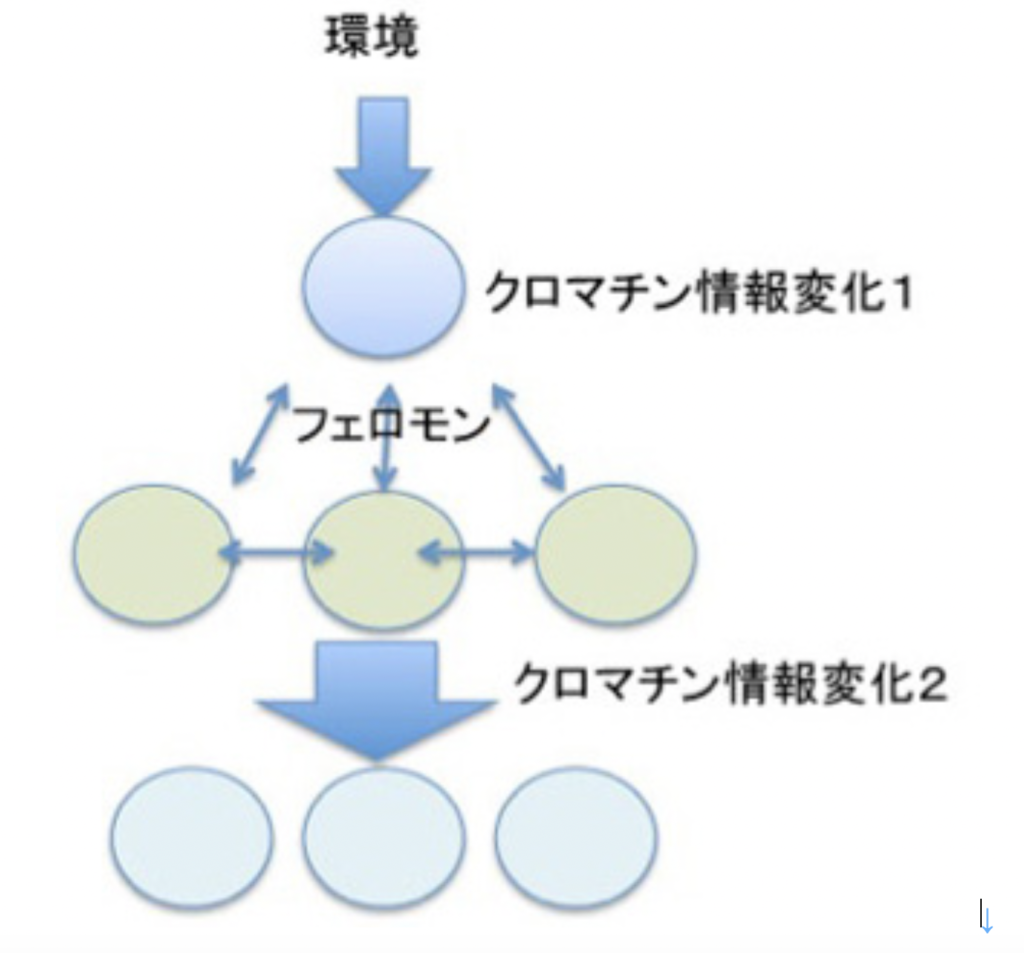
図4:環境ストレスによるクロマチン情報の書き換え機構にフェロモンが組み合わさると、より複雑な細胞間のコミュニケーションが可能になる。
図4で示した構造をよく見てもらうと、ある環境シグナルに対して直接対応する細胞と、その後フェロモンの作用で反応する細胞が別々に描かれているのに気づいてもらえたと思う。これは多細胞体制を想定して描いている。多細胞体制では、個体と環境、個々の細胞と環境、個々の細胞間、そして個体と個々の細胞間の4種類の関係が成立しており、この関係から生じる細胞の変化はすべてゲノムの変化を伴わない、クロマチン情報の変化が背景にある。例えば、個体と個々の細胞の関係には、同時的な情報の共有と、細胞レベルの経験の記録が必須で、これはクロマチン情報とフェロモン情報(細胞間情報伝達機構)だけで行われている。逆に多細胞体制成立から考えると、フェロモン情報とクロマチン構造を介する情報が誕生し、それが統合されることが、多細胞体制誕生の必須の条件であることになる。
[ 西川 伸一 ]
新しい情報と進化
2016年10月17日
前回まで、ゲノム、フェロモン、クロマチン構造が媒介する情報と進化で生まれた新しい情報について見てきた。これらの情報を、伝達、コミュニケーション、記録という3つの情報機能について見直してみよう。
進化についてのダーウィンの考えは、「集団は本来多様化する傾向を持っており、この多様化した個体の中から、環境に適合した個体が選択される」というものだ。ただ当時は全く理由の分からなかった個体の多様化の背景には、形質(フェノタイプ)よりはるかに大きな、ゲノムレベルでの多様化がある。
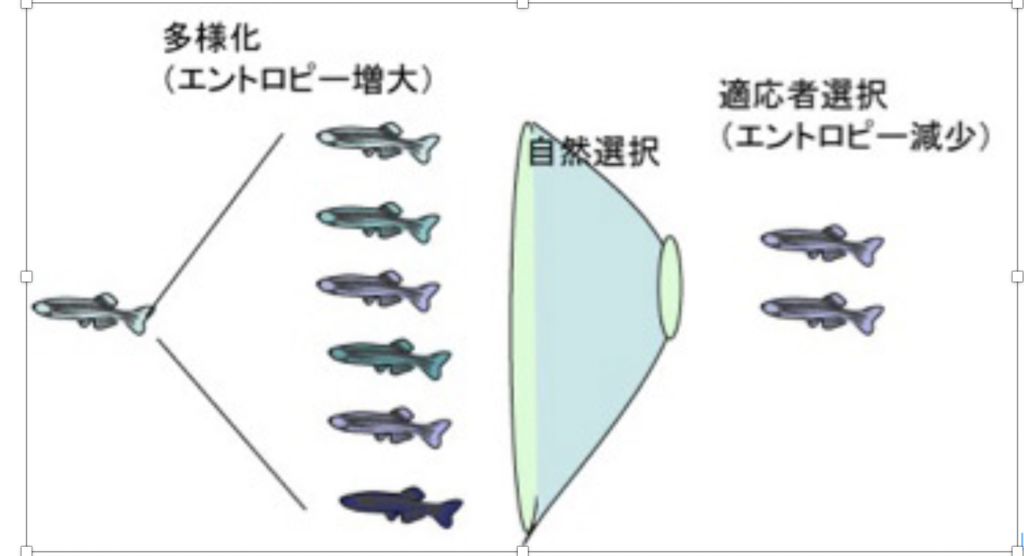
図1 ゲノムの多様化と自然選択だけから考える進化過程についての一つの表現。
ゲノム情報の多様化はエントロピーの増大と同じで物理的に自然におこる。この多様化したゲノムに対応する形質が環境にフィットした個体が選択されることになるが、これにより多様性(エントロピー)は、低下する。
この過程を単純な図式にすると図1のようになるが、この過程がただ続くとすると、せっかく獲得したゲノムの多様化も、選択により打ち消されるため生物が多様化する方向には進まないことになってしまう。
実際、全球氷結や隕石衝突など、生命自体にとって極端な条件が生まれると、ほとんどの種が絶滅し、生命の多様性は極端に低下する。ただ、通常はこのような条件が生まれるのは稀だ。なぜなら、地球は広く多様な環境が存在しているため、環境の違いだけ、それを同化した生物も多様化することになる。
ただこの説明のほとんどは、生命の情報としてゲノムしか想定していない。しかし私自身は、情報であれば全て進化に関わるものとして扱おうと考えて、これまで生命から生まれた全ての情報をリストする作業を進めてきた。もしフェロモン及びクロマチン構造を媒体とする情報のセットを生命の情報として認めるなら、当然この情報の多様化を統合して生物進化を考える必要がある。
そこでまず、フェロモンとクロマチン情報が新しい情報として加わることで、進化過程がどう変わるのか見てみよう。ダーウィン進化で問題になる情報は遺伝可能な情報であるという縛りがかかる。だとするとこの条件を満たす情報はフェロモンではなくクロマチン情報になり、クロマチン情報だけがダーウィン進化の駆動力として働く資格があるように見える。
しかし多様化できる情報をゲノムに限定しない新しい進化過程を構想するとき、フェロモンは記録情報でないからといって無視してもいいかというと、答えはノーだ。実際にはフェロモンにより新しい能力を獲得した集団が、環境を書き換えることで、能力の効果を記録してしまうことがありうる。例えば、古い旧家で何代もの子供が育ったとすると、先祖の建てた家という環境が、そこで育った子供たちに繰り返し制約を加える。この制約による変化も、ある意味で遺伝していると言って良い。
単細胞動物でも、多細胞動物でも、フェロモンは個体同士、細胞同士の行動の統合を可能にする仕組みだ。例えばクオラムセンシングを考えてみよう。この仕組みはフェロモンの濃度を感じて、周りに集る個体の濃度を感じ、十分な個体が集まったときに環境へ働きかけ、環境を自らの生存に適すように作り変えるのに使われている。このように、フェロモンに記録機能はなくても、環境側を変化させることで子孫へと一種の記録を残すことができる。この、環境を変化させるという記録の仕方は、後で神経細胞による情報を考えるとき重要になる(図2)。
他に、フェロモンは個体に働きかけ、接合を誘導したり、プラスミドを伝達したり、様々な方法で、ゲノムの多様化を促進する働きがある。こうして起こる多様化は、突然変異によるランダムな多様化ではなく、相同組み換えに基づくことで、多様化による生存への脅威を制約できるようになっている。すなわち、これまで生存を支えてきたゲノム情報の上に少しだけ変化を加える漸進的進化に適した変化の基盤になりうる。
一般的には、ダーウィン進化の駆動力、ゲノムの多様化は、DNA自体の化学的・生物学的性質により、自然に起こると考えられている。しかし、フェロモンが加わることで、変化させられた環境による新しい様式のゲノムや、クロマチン情報の選択が起こるようになる。また、接合誘導のような他の個体への働きかけの結果が、相同組み換えのような、単純なランダムなゲノムの多様化とは異なる、より制約を受けた様式による多様化が可能になる。このように、フェロモンだけでも、ゲノム多様化の様式を大きく変革できる(図2)。
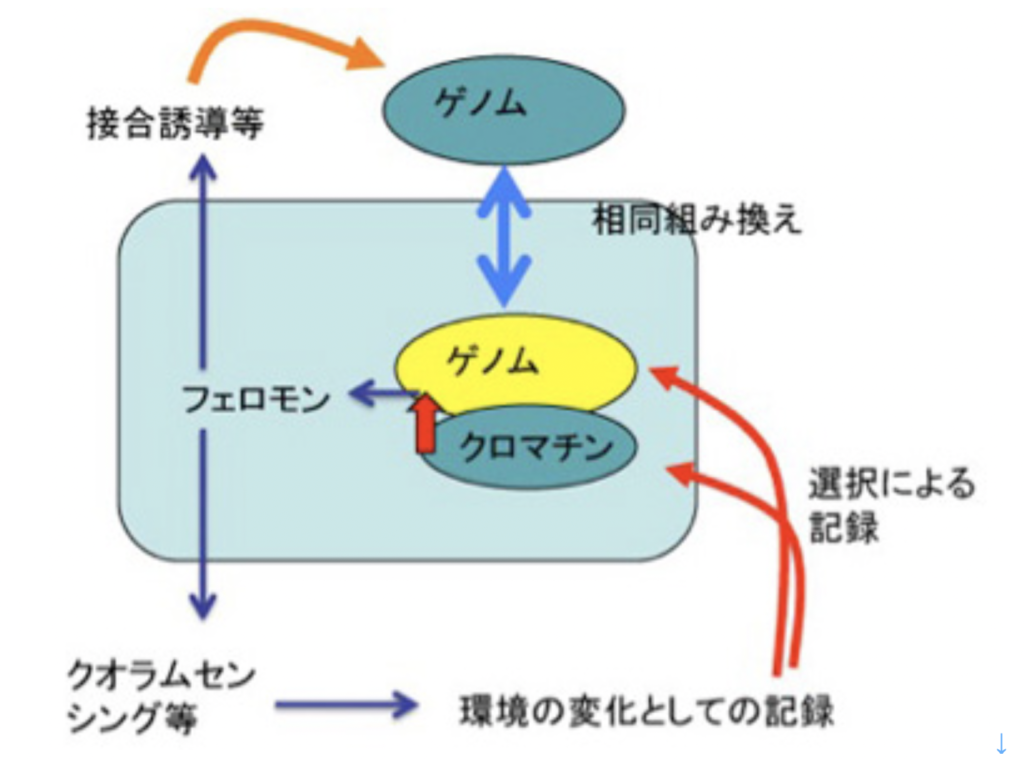
図2 フェロモンの影響は様々な形で記録できる。説明は文中
次に、記録できる新しい媒体クロマチン情報は、それ以前の進化過程をどのように変化させただろうか?
前回述べたように、ヒストン修飾やDNAのメチル化を介したクロマチン情報は、テロメアやセントロメアの形成を通してゲノムを構造化することに成功した。これにより、環状DNAでは限界があったゲノムサイズの増大が可能になる。そして、同じクロマチン構造変化を誘導するメカニズムが遺伝子転写のOn/Offを指示する情報へと発展する。このクロマチン情報によって、遺伝子発現パターンの調節、記録、そして子孫への伝達が可能になっている。
クロマチン情報で常に問題になるのが、「エピジェネティックな記録は子孫に伝わるのか?」という遺伝の問題だ。しかし、多くの細胞は分裂を繰り返しても、同じクロマチン情報を維持できる機構を有しており、原理的にはクロマチン情報は遺伝できると考えていい。
もちろん多くの動物の初期発生過程では、卵子と精子などの配偶子が持っていたクロマチン情報が一旦キャンセルされたあと、体細胞とともに生殖細胞が新たに発生する。従って、新しい個体が発生する過程では、親の持っていたクロマチン情報は消えることが多い。ただ、iPSリプログラムによるクロマチン情報の変化についての研究からわかったように、クロマチン情報を完全にリセットできないケースがある。このような場合、エピジェネティックな状態が子孫に伝わったとして、大きく取り上げられることになる。
いずれにせよ、単細胞生物や、あるいは培養細胞のクロマチン情報を調べた研究から、細胞分裂を経てもクロマチン情報は維持され、子孫に伝達されるのが普通だ。したがって、単細胞動物の進化過程では、クロマチン情報も子孫に伝達される情報として、進化過程の駆動力として貢献できたはずだ。
次にクロマチン情報が加わると、どう進化は変わりうるだろう?
- ①クロマチンによるゲノムの構造化により、ゲノムサイズや複雑さを格段に高めることに成功した。ただ、サイズの大きい複雑なゲノムになるほど不安定性は増し、またゲノムに大きな変化が起こることは当然個体の生存にとっては大きな脅威になる。しかし、クロマチン構造変化を介する情報が利用できると、ゲノムの変化に伴う個体への影響を減らすことができる。
このことは、ゲノムへ外来遺伝子が侵入してきた時、外来遺伝子の活性をクロマチン情報が抑える過程を見るとよく理解できる。われわれの細胞にレトロウイルスが感染すると、ウイルスゲノムの活性により宿主の遺伝子発現が大きく変化する可能性がある。しかし多くの場合、DNAのメチル化とヒストンのH3K9me3型への転換が飛び込んだレトロウイルスの周りで進み、その活性を抑えてしまう。
すなわち、飛び込んだ遺伝子はサイレンスされて個体に影響を及ぼすことはないが、ゲノム自体は多様化しており、ポテンシャルは上昇したといえる。すなわち、クロマチン情報により、遺伝子発現パターンが大きく影響されるのを食い止めながら、ゲノムサイズを増大させ、複雑化することが可能になる。
一時期、中立変異がどの程度可能かについて盛んに議論が行われたが、このときクロマチン情報の可能性については考慮されないまま議論が進められた。しかし、クロマチン情報によるゲノムの構造化は、ゲノムの変化が直接形質変化に反映されないよう、緩衝材の役割を持っていることがわかる (図3−①)。 - ②クロマチン情報は、環境ストレスやフェロモンシグナルに対して、多くの個体が同時に一定の反応を起こし、新しい状況に適応することを可能にする。例えば細胞増殖速度を落とし、あるいは環境変化に強い細胞を形成するなどがそうだ。この環境への早い一様の対応により、クロマチン情報誕生前はゲノム情報がそのまま形質に反映し、その適合性で個体自体選択されていた状況は大きく変化する。すなわち、様々な環境変化やシグナルに対し、クロマチンシグナルによりまず多くの個体が共通の反応を起こし適応する。このクロマチン情報レベルの変化は、環境に適応した個体数を増やし、またゲノム内で変化しやすい部位とそうでない部位の違いを生み出す。選択を受ける個体数が多いと、選択後の個体数の増加が望める。また、変異が選択的に起こることで、生存への選択圧を下げたり、あるいは上げたりしてより環境に適合(選択圧を下げた)した状態でのダーウィン進化が可能になる。
- ③あまり議論しなかったが、フェロモンとクロマチン情報は多細胞体制誕生に必須の条件だ。多細胞生物では、フェロモンを起源とする様々な細胞間シグナル伝達分子により、細胞のアイデンティティーが決められるが、アイデンティティーの確立には全てクロマチン情報の書き換えが必要になる。もちろん、多細胞体制への進化には他にも様々な条件が必要だが、細胞間シグナル伝達と、クロマチン情報は中でも基本中の基本条件になっている。こうして多細胞体制が生まれると、環境と個体との相互作用のあり方が大変革する。すなわち、個体の異なる構成部分は環境と異なる様式で関わる。例えば、葉で太陽を利用し、根は地下で水を利用する。この環境との相互作用の様式の大変革が、進化の様式を多様にし、例えばカンブリアの大爆発をもたらすことになる。
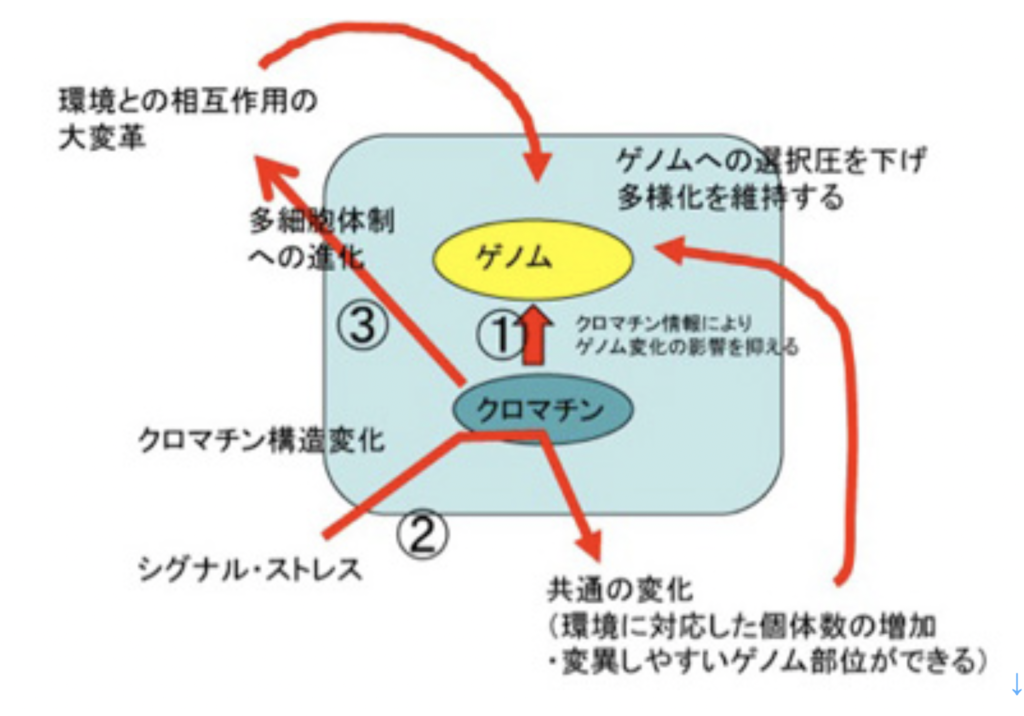
クロマチン情報の誕生による、進化様式の変化。説明は文中。
以上、クロマチン情報とフェロモンが情報に加わることで、進化の様式が大きく変革することを理解してもらえたのではないだろうか(図3)。
これまで、進化の様式が大きく変化するためには、選択圧が高まることの必要性が強調されることが多かった。しかし、情報を統合的に考えることで、私は逆に淘汰圧が下がり、最終的に種の多様化を決めるゲノムの多様化が保障されることが、生命の多様化には重要でないかと思っている。これまで述べてきたように、ダーウィン進化の結果は環境の自己への同化だ。多くの環境要因を同化できるということは、種が多様化することと同じだ。図1に示したゲノムしか情報がない場合の単純な進化過程をいくら積み重ねても、真核生物誕生後の急速な進化を説明することはできないのではないだろうか。ただ残念ながら、これは私の個人的意見に過ぎないと断っておく。
ゲノム、フェロモンからクロマチン情報へと進んだ情報の多様化と統合過程の繰り返しが、次は神経細胞の誕生から始まる。
[ 西川 伸一 ]
動物と神経の誕生
2016年11月1日
神経系がゲノムやクロマチン情報とは全く異なるレベルの情報システムを形成していることは明らかだ。ただ、これは神経系が高度に発達した動物についての話で、神経細胞誕生=新しい情報の誕生ではない。即ち、無生物から生物が誕生した過程と同じ問題が、神経系から新しい情報が生まれる過程に存在している。そこで、今回から神経細胞の誕生によって可能になった情報システムの成立について、動物進化過程を振り返りながら考えようと思っている。
この新しい情報システムは、もちろんゲノムやクロマチン情報とは密接な相互依存関係にあるものの、半独立の情報システムを形成していると言える。
例をあげて説明しよう。これまで述べてきたフェロモン、クロマチン情報は、ゲノム情報から独立して情報として働くことはできない。例えば、クロマチン情報はゲノムを構造化するとともに、この構造を変化させることでゲノム上の様々な大きさの領域にある遺伝子が利用可能かどうかを決定する(65話図3)。すなわち、クロマチン情報はゲノムとの関係を前提に複製可能で、その情報はゲノム上の遺伝子発現の違いとして表現される。誤解を恐れず例えるなら、ゲノム上の遺伝子の発現を指令するアルゴリズムと言える。
同様にフェロモンも、クロマチン情報やゲノム情報の変化を誘導することが主要な機能だ。フェロモンやクロマチン情報は独自の情報として振舞っても、ゲノムから決して独立することはない。
しかし、神経系による情報の解釈、生成、伝達は、ゲノムは言うに及ばず、クロマチン情報からもかなりの程度独立していると考えられている。実際、人間の脳が生み出した情報は、ゲノムやクロマチン情報からは独立している。だからこそ、神経系をコンピュータで再現できると考える科学者は多いし、また脳自体をコンピュータの一種として理解しようとする科学者は多い。
私自身は、神経を媒体とする情報システムは、コンピュータと同じで、神経細胞同士が階層的なネットワークや回路を基盤にしたシステムだが、その回路自体がクロマチン情報やゲノム情報と相互作用できることが、神経系独自の情報システム形成に重要な役割を果たしていると思っている。ただ、この問題については最初から議論するのを避けて、まず神経細胞の誕生から脳の誕生に至る神経系の進化を段階的に見ていきながら、それぞれの段階で神経系を情報の観点から眺め直すとともに、結合性とクロマチン情報や、ゲノムなどの下位の情報との相互作用について考える。
最初は神経細胞の誕生だ。現存の生物の系統樹から判断すると、神経細胞は動物、すなわち動く能力を持った多細胞生物の誕生とともに生まれてきたことは間違いない。図1に示した系統樹では現存の多細胞生物を5種類に大別しているが、海綿動物とセンモウヒラムシには、いわゆる神経細胞は存在しないことがわかっているが、残りの動物は全て神経細胞を持っている。
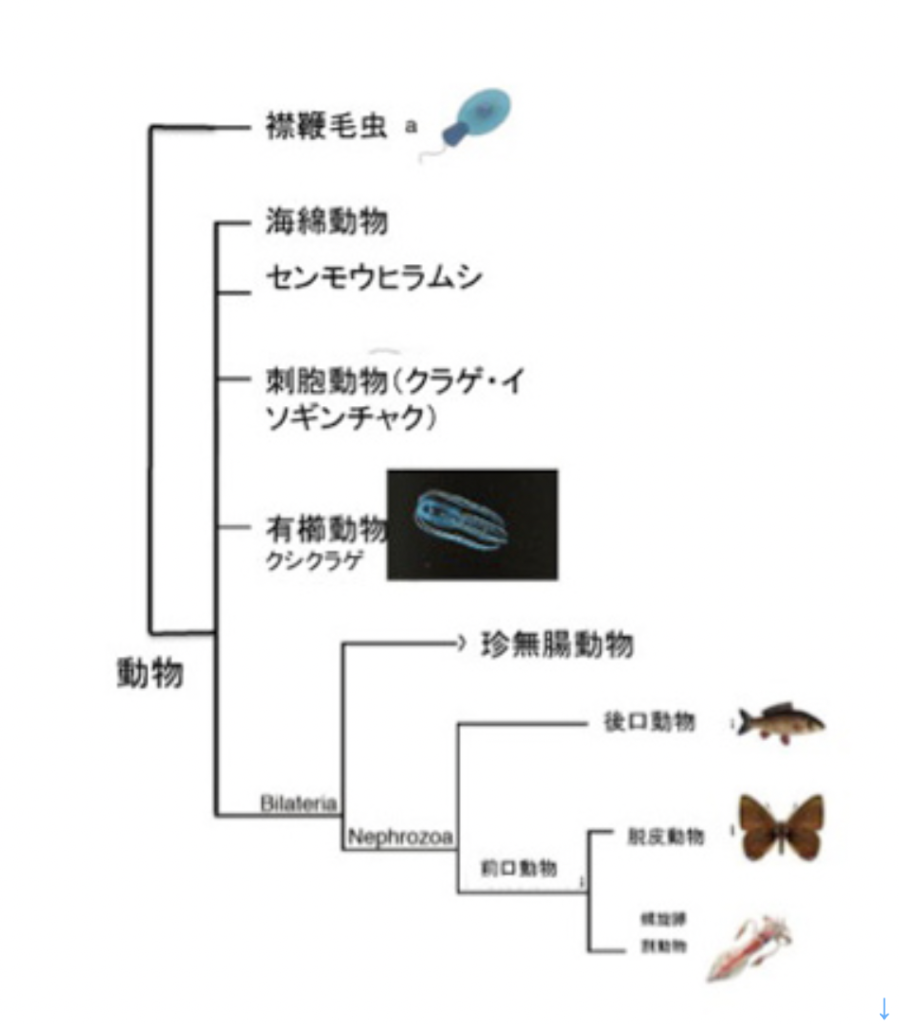
図1:動物の系統樹 現存動物の系統樹。この図では、神経細胞や筋肉細胞を持たない海綿やセンモウヒラムシを、他の動物から区別して表しているが、ゲノムから得られる系統樹とは一致しない。
では神経細胞は動物が進化した後現れたのか?
海綿やヒラムシに最初から神経がなかったかどうかは議論が分かれている。というのも、図1のクシクラゲは、ゲノム系統樹から見ると他の4種の動物から最も早く分離した系統で、左右相称動物から見て系統上最も離れている。ところがこのクシクラゲに神経細胞が存在することから、より左右相称動物に近いヒラムシや海綿にも最初は神経細胞が存在し、その後神経細胞を退化させたと考える説もある。
これとは逆に、神経細胞と、他の興奮性の細胞とを区別する指標として用いられるナトリウムチャンネルは左右相称動物だけにしか見つからないことから、クシクラゲと他の動物が別れた後、クシクラゲと左右相称動物は独自に神経細胞を発生させたと考える説もある。
いずれにせよ、最初の神経は私たちが一般的に持っている軸索、細胞体、樹状突起からなる典型的な神経ではなく、外界からの刺激に反応し、その興奮を他の細胞に伝達する能力を備えていても、より普通の細胞に近い形態を持っていたのではないだろうか。実際、興奮性の細胞系列の一部は、興奮を力に変える筋肉細胞へと発展する。一方、様々な刺激を感知して、他の細胞に伝達する能力を持つ細胞から神経細胞が生まれたのだろう。細胞の興奮に必要なイオン勾配の維持機能、そのイオンを選択的に通過させ膜電位を発生させるイオンチャンネル、そして興奮を他の細胞へ伝える化学システムは動物の誕生前に進化し、採用を待つだけになっていた。
では、神経細胞が生まれることで何が可能になったのか、現存の生物で見てみよう。
図2にゴカイの幼生の光受容体神経細胞について示すが、環形動物幼生の神経系は、まだ組織化されない単独の神経細胞が、どう神経細胞進化の初期に使われていたかを考えるいい材料になる(図2)。
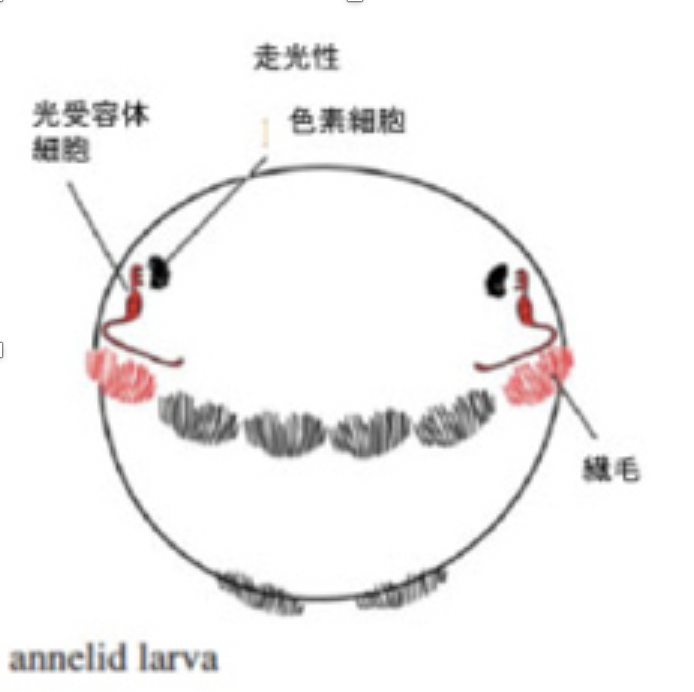
図2:ゴカイ幼生の走化性
G.Jekely et al : An option space for early neural evolution,
Pil.Trans.R.Soc.B :
370:2015.0181 Fig2Aより改変、この図の元となったoriginal 論文はJekely et al, Nature 456, 395, 2008)
ゴカイの幼生は繊毛を使って水中を浮遊しているが、光を感じると繊毛運動を調整して光の方向へ移動する走光性を示す。これに関わるのが図2で示した3種類の細胞、色素細胞、神経細胞(光受容体細胞)、そして繊毛を持つ上皮細胞だ(original 論文はJekely et al, Nature 456, 395, 2008)。図に示す様に色素細胞とそれに結合する神経細胞により色素細胞で吸収された光エネルギーが神経興奮として受容され、同じ神経細胞が繊毛上皮とコリン作動性のコンタクトを形成することで繊毛の動きを調節している。片方の目にだけ光を当てると、光を受けた方だけ繊毛の運動頻度が低下し、結果幼生は光の方に進む。
この研究から、神経細胞の出現により、外界の物理的刺激を感知して、他の細胞(近くの繊毛上皮細胞)にそれを伝達して、活性を変化させる一連の過程が可能になっているのがわかる。
光や温度、あるいは圧力などの物理変化に素早く対応することは、生物の生存にとって重要な条件だが、神経細胞出現後の素早い反応はフェロモンのような化学的シグナル分子だけでは実現できない。このため、光などの外界の物理的刺激に素早く反応するための分子メカニズムは動物誕生以前から存在していた。例えば、単細胞生物クラミドモナスの走化性に利用される光を感じて開くイオンチャンネル、チャンネルロドプシンはその典型で、この分子を使った光遺伝学は、脳研究を席巻する勢いだ。
このような様々な物理刺激を感知して素早く個体の行動を変化させるシステムは、高等動物への進化過程で神経系の属性へと集約していくが、ゴカイの幼生の走光性行動には様々なメカニズムが組み合わさっている。
動物の運動のほとんどは筋肉、あるいは筋上皮と呼ばれる筋肉と上皮の中間のような細胞により行われる。実際、ともに興奮性の細胞と言える神経と筋肉は動物の誕生とほぼ同時に出現している。一方繊毛による運動は例えばゾウリムシなどの繊毛虫から見られるシステムで、より古い起源の運動生成システムと言える。私たち人間も含め、繊毛運動が神経支配を受けることは珍しく、その方向性やパターンは上皮の持つ極性を含むplanar polarityと称される特性により決められている。ゴカイの幼生は、この古いplanar polarity様式と神経支配という新しい様式を組み合わせて運動を調節している面白い例だと言える。
神経細胞により繊毛運動の強弱を直接神経支配することは難しい。結局ゴカイの幼生で見られるように、繊毛を持つ上皮自体に働いて、その細胞の持つ繊毛の運動を全て止めるという方法をとらざるをえない。
またもともと繊毛は早い大きな動きには向いていない。このため動物では、神経とアクトミオシン収縮系を備えた筋肉とがセットで働く”運動”が優勢になっていく。この組み合わせにより、一つの神経シグナルを複数の筋肉細胞の収縮に使うことが可能になり、大きな力を発生させることができるようになった。
もちろん、神経支配による上皮の活性調節は、ホルモン分泌の神経支配などとして今も見ることができる。
以上繰り返しになるが、ゴカイの幼生で見られる神経細胞の役割は、1)物理刺激を感受して、2)それを素早く他の細胞に伝達し、3)個体全体を刺激に対応させる、ことに尽きる。また同じ系が収縮性の筋肉細胞と組み合わさると、繊毛運動をはるかに超える大きな運動が可能になる。
最後に、神経細胞誕生により生まれた新しい可能性を情報と進化の観点から考えてみよう。
高等動物の脳神経システムには、間違いなく新しい情報システムが存在しているが、神経細胞や筋肉細胞の誕生により、これにつながる新しい情報システムが誕生したとは言えない。例えばクオラムセンシングでのフェロモンの働きに見られるのと同じ、環境の変化を感受して決まったアクションを起こす、インプット・アウトプットの関係が、より複雑化しただけと言っていい。事実、ナトリウムチャンネルの出現といった、新しい分子の誕生によるメカニズムの多様化がこの過程で起こったものの、例えば脊椎動物進化で抗体分子が現れるといった、本当のイノベーションは起こっていない。
しかし、物理刺激を感知できるようになり多様な環境変化に対応できるようになったこと、素早い反応が可能になったこと、そして筋肉の誕生により大きな運動が可能になったことは、環境を自己に同化する進化の駆動力の観点から見ると、大きなイノベーションだったと思える。
環境を感知して自己を守る反応という点ではフェロモンと同じだが、素早く、方向性を持った動きが可能になると、好ましくない環境を回避することで生存する個体数は当然増える。さらに、筋肉が生まれると素早い大きな回避行動をより刺激に合わせてコントロールすることで、多様な生息環境を選ぶことができる。これは環境を作り直したのと同じことで、環境の自己や種への同化が起こっていると見ることができる。こうしてより適した環境に移動することで、当然選択圧は下がり、多様性を維持する方向に働く。
このように、フェロモンも、神経や筋肉も、ゲノム情報による自然選択をできるだけ回避する方向に働く。実際、生きている個体の適応性は、それ以前のゲノム多様化と、選択の結果だ。新しい現在の環境に適合するかは試されたことがない。これに対し、新しく生まれた情報システムは全て、今生きている個体の生存に関わる。このように、今という時間への適応を拡大することが、生物の多様化に貢献したと思う。神経や筋肉で言えば、運動性が質及び量の面で大きく高まることで、動物は急速に地球上の様々な環境へと拡大できた。すなわち、過去の進化で獲得された環境適合性に合うよう個体が環境を選び、場合によっては改変することで、選択をかわしてゲノムをさらに多様化できたと思う。
新しい情報誕生まではまだまだだが、神経細胞の誕生はさらに新しい展開を見せる。次は記憶の誕生について見ていく。
[ 西川 伸一 ]
神経記憶 I
2016年11月15日
今回から記憶について考える。
記憶自体は情報ではない、しかし、皆さんのパソコンでの情報処理に記憶素子が欠かせないように、情報が成立するために欠かすことができない機能だ。情報を論じるために記憶の問題を避けて通れないことを理解してもらうため、これまで見てきた情報を、記憶という観点から見直すところから始める。
通常「記憶」という言葉は、経験によって脳内に誘導された神経活動パターンを、時間を経た後で呼び起こすことを意味する言葉として使っている。この神経記憶については後に考えるとして、この言葉が私たちの神的活動にとどまらず、あるパターンを繰り返し呼び起こす現象を表現するのに使われていることを思い出して欲しい。例えば、形状記憶合金というと、形が壊れてもまた自然に同じ形を取り戻すことができる合金を意味している。
生物学でも記憶という言葉が使われる現象は多い。例えば免疫記憶は、一度経験した抗原に対して免疫系が最初よりより速やかに、しかもより強い反応を起こす過程と定義できる。免疫記憶の背景には、かなり複雑な細胞間相互作用が存在するが、極めて単純化して説明すると、経験した抗原に反応するリンパ球が選択的に増加する、あるいは寿命が長くなることにより記憶が形成される。すなわち、経験を、この抗原特異的リンパ球の数や活性の変化に変換して記録している。
このように何らかの形で一定のパターンを記録することを記憶と考えると、生物にとって最も根源的な記録を担うDNAも一種の記憶と呼ぶことができる。実際、ゲノムは、ゲノム自身の複製だけでなく、それを持つ生物の複製も可能にしている。そして、複製とは同じパターンが繰り返すことに他ならない。もし、コンピュータのメモリーのように記憶=記録情報だとすると、ゲノムをメモリーと呼んでよさそうに思える。
しかし、塩基配列としてゲノムに書かれている情報を決して記憶と呼ぶことはできない。なぜなら、ゲノム複製も、細胞複製も、ゲノム情報だけでは実現できず、ゲノムの情報に基づいてオペレーションを行うための様々な条件が揃って初めて可能になる。以前、マイコプラズマの再構成実験を例に(http://www.brh.co.jp/communication/shinka/2016/post_000013.html)この問題について議論したが、今の所ゲノムは生きている生物にしか移植できない。同じことは、発生初期の受精卵では胚自体のゲノムからの転写がほとんどないことからもわかる。個体の一生を通じてあらゆる細胞で働くことができるゲノムも、まず母親から受け継いだRNAから作られた分子による発生のプログラムが活性化されて初めて、その個体の情報として働き始める。
このように、記憶は記録された情報ではなく、体験、記録、呼び起しという順序で進む一連の過程全体に他ならない。例えば、図1に示すように、ゲノムは情報として二つの役割を担っている。
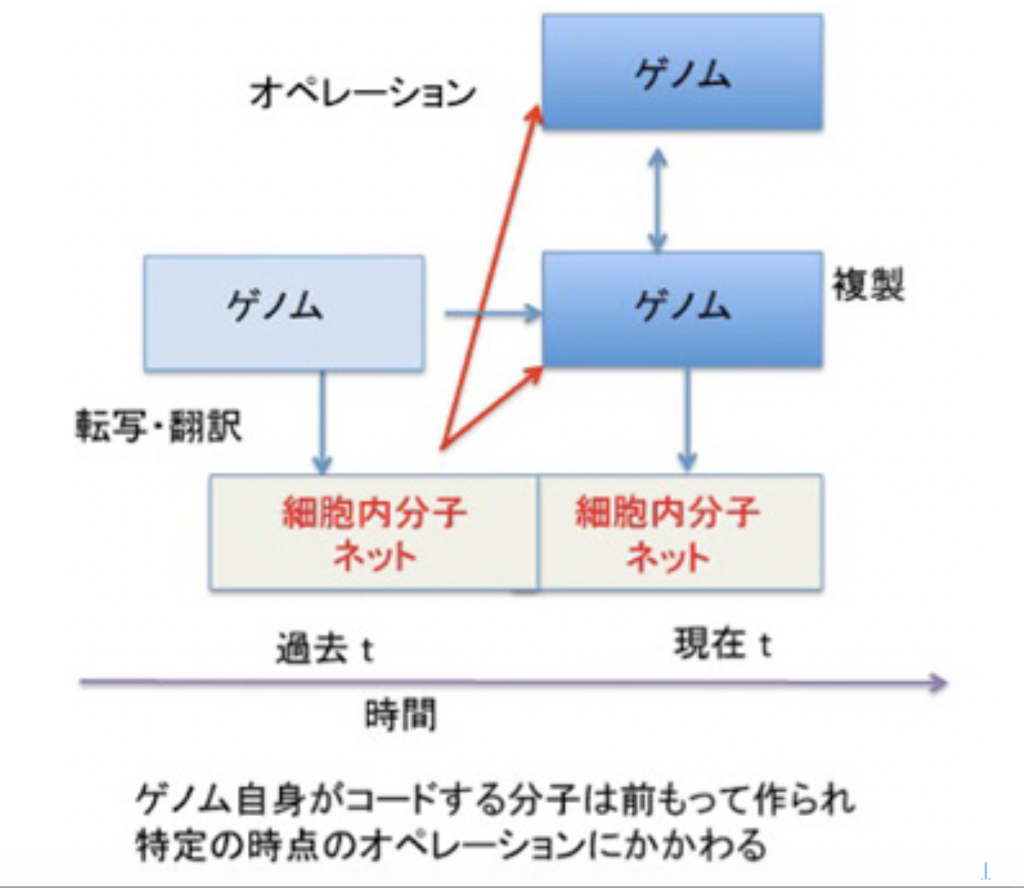
図1 ゲノムは生物の持つ最も基本的記憶過程と考えられる。説明は文中
一つは自分を複製するための情報と、複製のための鋳型としての役目だ。ただ、ある特定の時点(図1:現在 t)のゲノムのオペレーションは、同じゲノムによりその時点より前に作られた(過去 t)分子ネットワークによりオペレートされている。生命特有の鶏が先か、卵が先かという問題がここでも現れる。いくら外来遺伝子を細胞の中で機能させられるからといって、ゲノムとそのオペレーションは互いに分離不可能で、ゲノム=記録として切り離すことはできない。
ではなぜ今使っているコンピュータでは、アプリケーションや記録をメモリーに記録して分離することができるのか。これは、コンピュータ自体が一定の命令で動くように人間により作られているからというしかない。生命に、コンピュータにとっての人間のように、明確なデザインの出し手は存在しない。
ゲノムのオペレーションがそれ以前に準備された分子ネットワークに依存しているとすると、ゲノムのオペレーションに関わる分子ネットワークを辿れば、結局最初の生命誕生時にゲノムをオペレーションした分子ネットワークに行き着く。すなわち、ゲノムが38億年前に誕生した最初のゲノムまで連続しているのと同じで、細胞内の分子ネットワークも途切れることなく過去から現在そして未来と、ゲノムとともに受け継がれる。このように、現存の生物の記憶には無限の過去の経験が持ち込まれていることになる。こう考えると、進化自体DNAを記録媒体とした記憶と考えることができる。言いかえよう。ゲノムを記憶という観点から考えると、必然的に38億年という全進化史が含まれる壮大な過程と見ることができる。どの体験がゲノムに記憶され現存の種に維持されているのかを研究するは、進化学に他ならない。
一方、クロマチン情報は、基本的には各生物の一生という時間枠の中での記憶と言える。この意味で、神経記憶と同じだが、クロマチン情報が記憶と定義することに抵抗を感じる人も多いだろう。ただ思い出して欲しい。生物発生では、卵から個体誕生まで、ほぼ同じ過程が概ね正確に繰り返される。この過程では、ゲノムは変化せず、時間、場所に応じてクロマチン情報が変化していく。例えば発生学でよく研究される両生類の初期発生を例に見てみると、どの場所に精子が入るかは偶然の結果だが、侵入点が決まると体の大まかな体制が決まるよう、すでに卵細胞のゲノム、クロマチン情報、分子ネットワークが準備されている。
核移植クローン実験から明らかになったように、卵細胞質の分子ネットワークの多能性プログラムを維持する能力は高い。しかし、この分子ネットワークは卵割に伴い内外から新しい刺激を受けて分子ネットワークを変化させると共に、次々と生まれてくる細胞のクロマチン情報を変化させて、分化可能性が制限された分化細胞を形成する。一旦分化細胞のクロマチン情報が確立すると、分化した細胞の性質は安定に維持される。
クローニングやiPSにより示されたように、分化した細胞のクロマチン情報による分化制限を元に戻すことはできないわけではないが、普通はこの制限のおかげで、安定に同じ運命の細胞を作ることができる。こうしてそれぞれの細胞でクロマチン情報を中心に新しい記憶システムが成立する。
このクロマチン情報を中心とする記憶システムは、プログラム前の分子ネットワークと、外から入ってくる新たなシグナルにより少しづつ変化するが、このオペレーションメカニズムについては研究が始まったばかりでわかっていないことが多い。ただ最近、比較的少数の細胞でゲノム全体をカバーするクロマチン情報を調べる手法が開発されているので、急速な研究発展が期待できる。
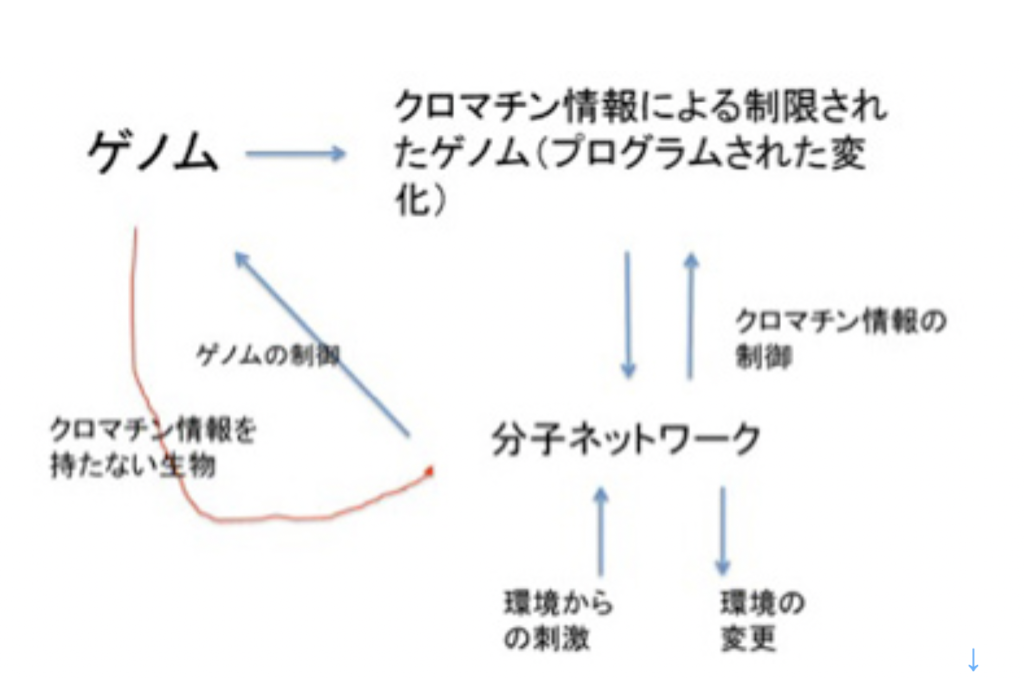
図2:クロマチン情報を中心とする記憶システム。説明文中
ゲノム、クロマチン情報、分子ネットワーク、外界からの刺激などを記憶の観点から見た関係を図2に示す。ここでは、ヒストン修復と、DNAのメチル化の制御によりクロマチン情報が書き込まれることで、特定の細胞を繰り返し作るための長期記憶が形成される。この記憶=クロマチン構造の維持と変更システム、により制限を受けた転写により生まれる分子ネットワークが、分化した細胞のオペレーションの基盤になる。各細胞の分子ネットワークは内外からの(周りの細胞も含む)様々な刺激により変化する。この変化は、一種の短期記憶と見ることができるが、クロマチン情報の制限を外せない限り、長期記憶にはなりえない。ところがある特定の刺激が分子ネットワーク内に揃うと、ヒストン修飾やDNAメチル化の変化による、クロマチン情報の書き換えが起こる。これにより、細胞の分子ネットワークの変化として短期的に維持されていた記憶が、より長い期間維持される長期記憶に変化する。
ある特定の細胞のクロマチン情報の書き換えを可能にする分子ネットワークの変化について研究しているのが、リプログラム研究で、山中4因子の発見に端を発して、様々なリプログラミング、すなわちクロマチン情報書き換えのための条件が明らかになりつつある。
神経記憶から脱線したが、図1、2に示した記憶システムは、真核生物のあらゆる細胞で見られる。従って、神経細胞記憶もこの2つの記憶システムの上に発展したといえるが、これまでの記憶システムとは大きく違っている点がある。すなわち神経記憶は、動物の誕生とともに新しく生まれた興奮性の神経細胞同士の結合により(ここでは、神経細胞を、同じ前駆細胞から生まれるニューロン、アストロサイト、オリゴデンドロサイトを全て含む広い定義を用いる)新たな記憶システムを成立させている点だ。
この新たな神経記憶の特徴については次回から考えるが、最後に神経記憶もクロマチン情報を中心とする記憶システムの上に構築されていることを、前回に例として使ったゴカイの幼虫の神経系をモデルに見ておく。このモデルを選んだのは、これが一本の神経細胞だけで形成されている最も単純な神経系で、なおかつ神経細胞の基本的特徴を備えているからだ。そこで、ちょっと想像をたくましくして、この系での記憶について考えてみた。もちろんこれから述べることは証拠があるわけではなく、神経記憶自体は回路形成なしでも考えられることを知ってもらうために考えたたとえ話だ。
さて前回述べたように、ゴカイ幼虫には、光をトラップする色素細胞と協調して光に反応し、その興奮を繊毛細胞に伝え、繊毛の動きを止める神経細胞がある(前回図2、および今回図3)。この神経は感覚神経と運動神経の両方の役割を持つため、神経回路は形成しない。
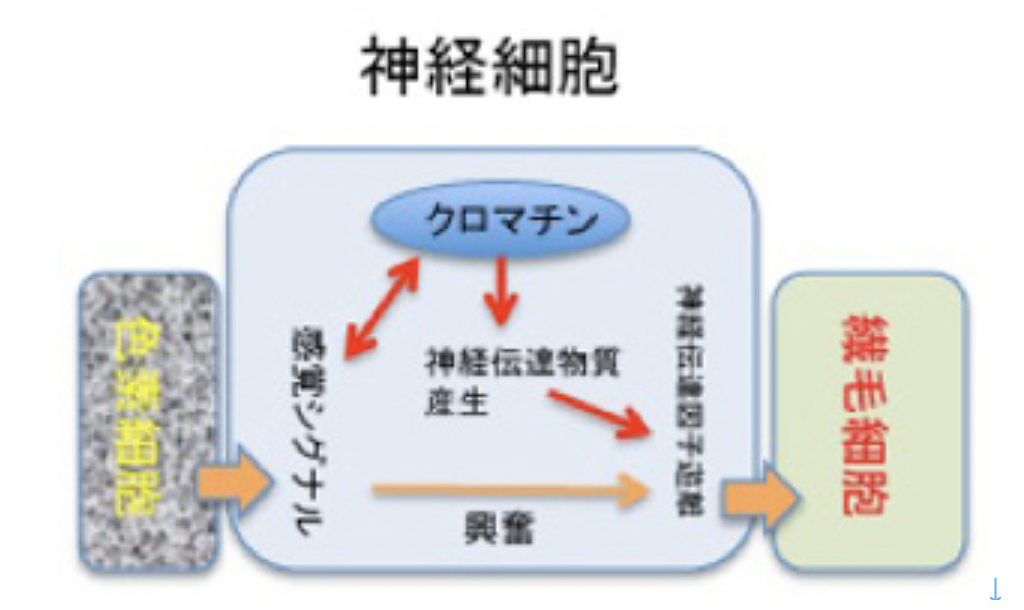
図3:ゴカイ幼生の感覚運動神経システム 説明文中
この系では神経細胞が感覚刺激を感受すると、チャンネルが開き細胞が興奮、その興奮によりアセチルコリンが遊離、繊毛細胞に働いて繊毛の動きを止める。これは一般的神経細胞の興奮と同じで、分子ネットワークに大きな変化はない。ただ、この状況でも光刺激が繰り返すとアセチルコリンが枯渇する。このため新たな合成が始まるまで、幼生の運動は停止する。この結果、幼生はストップ・アンド・ゴーを一定のサイクルで繰り返す。
この時、光シグナルの強さによりもしクロマチン情報が変化し、アセチルコリンの合成速度が完全に変化したとすると、ストップ・アンド・ゴーの感覚を長期的に長くしたり、短くしたりすることができる。即ち、刺激による長期記憶が誕生したことになる。
繰り返すが、これは理論的可能性を述べただけで、実際にどうかはまったく知らないが、神経内の分子ネットワークとクロマチン情報を変化させることで、回路形成なしに神経記憶を形成することができることを理解してもらえたと思う。
この図1,2の記憶システムの基盤の上に新たに回路形成が加わったのが神経記憶で、次回はこの回路形成が加わったことによって生まれる新しい可能性について見ていく。
[ 西川 伸一 ]
神経記憶II
2016年12月1日
前回まで、ゲノム情報を中核とする記憶(Geneme mediated memory:GMと呼ぶ)、クロマチン情報を中核とする記憶(Chromatin mediated memory: CMと呼ぶ)について考えを述べてきた。GMは生命進化全体、CMは生物個体の発生から死までの時間をカバーする記憶システムと言える。
生物学は現象を生物の側から説明しようとして目的論に陥ることが多いが、それを承知で言うと、GMもCMも、過去の経験を記憶し、未来に備えるシステムと言える。
前回見たように、GM、CMそれぞれの記憶システムは階層的に重なって統合されており (前回:図2)、CMはGMを基礎としてその上に形成される。この構造はCMを有する全ての真核細胞で共通で、当然神経細胞も他の細胞と同じように、GM,CMを持っている。このおかげで単一の神経細胞だけでもCMを利用して神経記憶反応を起こせる可能性を前回示した。他の細胞と比べると、神経細胞は刺激に対して早い反応(興奮)を起こすことができるため、神経記憶に見えるが、記憶自体は神経特有というよりCMの機能を反映している。
このGM、CM記憶の上に新たに生まれたのが、神経細胞同士の結合した神経回路形を中心にした記憶(neuron mediated memory:NM)で、これにより動物は、質的、量的に一段高い次元の記憶システムを獲得し、今や地球上に100億個体も存在するという驚くべき繁栄を遂げた人間を進化させる大きな原動力となった。
GM,CMでは、ゲノム情報やクロマチン情報のように分子レベルの媒体が経験の記録を担っているが、NMは細胞間に形成される回路のように細胞レベルの媒体を利用する記憶と言える。神経細胞の特徴は、細胞同士で刺激や抑制の回路を形成できることで、この回路が経験の記録の単位になる。もちろん細胞間のネットワーク形成が見られるのは神経だけではない。例えば、免疫細胞記憶成立には、神経伝達と同じように、サイトカインと呼ばれる分子による、シグナル伝達機構が存在する。また、一つの免疫反応に関わる細胞の種類は、マクロファージや樹状細胞から、何種類かのT細胞、場合によってはB細胞と、神経系と同じように数が多い。しかし、免疫系が神経系と大きく異なる点は、抗原と、それに対する抗原受容体の間の特異的分子結合がネットワークの中心にある点だ。さらに、抗原が常に存在しないと、記憶も維持できないし、反応も起こらない。さらに、抗原刺激とそれを中心にする細胞間相互作用により、それぞれの細胞の増殖や寿命の延長などがおこるため、結果細胞の動態に大きな変化がおこる。
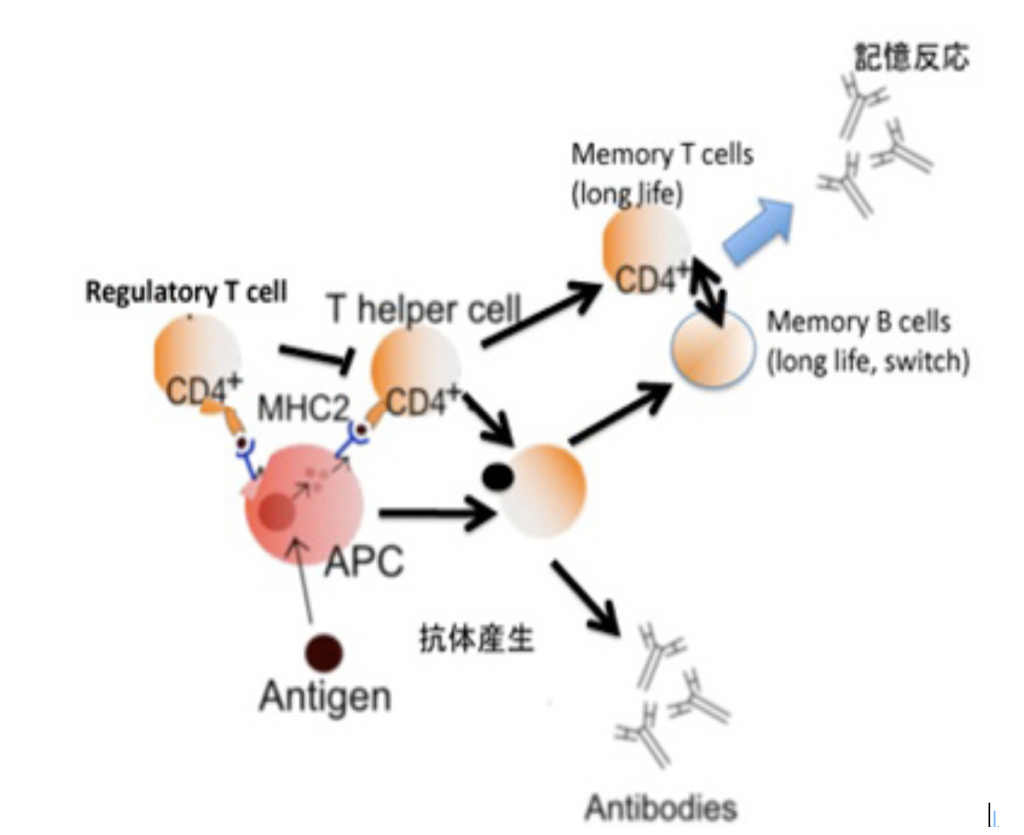
図1 抗体反応での免疫記憶の成立に関わる細胞間相互作用。
図には示せていないが、あらゆる細胞間相互作用に様々な形で抗原とそれを認識する細胞側の受容体が関わっている。
一方、神経細胞回路の場合、匂いや味の感覚のように、外来の化学物質により刺激誘導される場合でも、興奮が神経間で伝達されるときには、最初の興奮を誘導した物質は必要なくなっていることが多い。すなわち、感覚器の興奮が伝えられるとき、感覚を誘導した刺激は必要ない。もちろん刺激の持続時間が短い光などの物理的刺激は、最初から刺激の持続を前提に感覚を伝えることはできない。記憶が維持されるためにも、最初の刺激は必要ない。このように、神経記憶は刺激物質を中心に反応が構成されるのではなく、最初から感覚刺激を複数の神経細胞興奮のパターンへと転換された後、伝達され、記憶される。
どうして最初の刺激を誘導する物質とは全く無関係に、神経回路だけで、記憶が成立、保持できるのかを考えるのが、これからの問題で、詳しく見ていきたいと思う。ただ、神経科学についての私の知識は乏しい。このためこれから書く内容には、間違った記述が含まれる可能性があることを断っておく。
まず、神経細胞回路がどのように記憶に関わることができるか見ていこう。
まず個々の神経細胞をみると、一つの記憶素子として機能できることがわかる。図1に最も単純な電気的記憶素子DRAMを示すが、一個の神経細胞はDRAMに似ていると言える。図に示すようにDRAMは基本的にはスウィッチ役のトランジスタとコンデンサからできており、コンデンサーに電気を貯めることで1 or 0のデジタル記憶が可能だ。
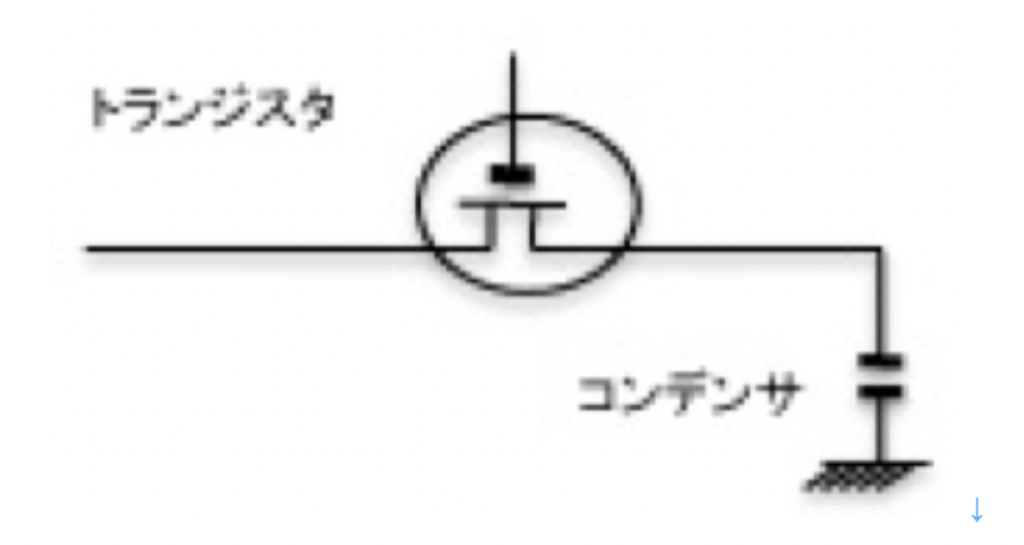
図2 記憶素子DRAMの構造:一般的にトランジスタは電子的スウィッチと考えればいい。
すなわち、スウィッチがオンになって電気がつながれば、コンデンサに電気が溜まった状態になる。一方、スウィッチがオフになると、コンデンサは自然放電して溜まった電気はなくなる。この溜まった電気があるときとないときで、2進法の1か0を記憶できる。
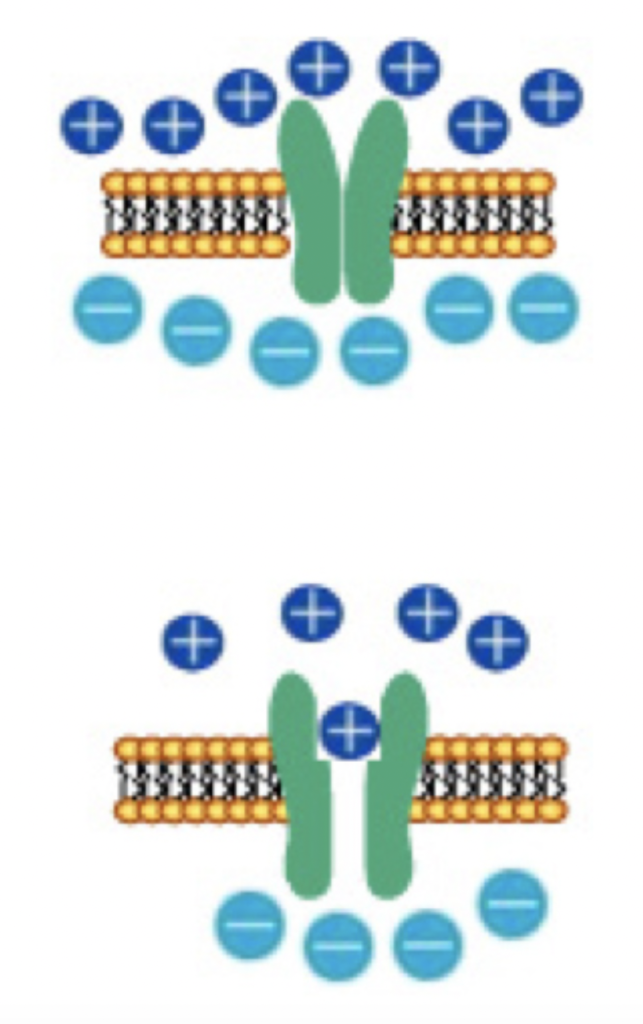
図3:細胞膜に形成された、イオン蓄積システムと、スウィッチ役のイオンチャンネル。
一方、神経細胞はイオンポンプを用いて外界と細胞内の様々なイオン勾配を保つことで、電圧を維持している。細胞膜には特定のイオン分子(例えばNaやK)を選択的に通過させるイオンチャンネルが存在し、刺激によりイオンチャンネルが開くと、特定のイオンだけを勾配に従って流すことができる。すなわち、イオンチャンネルはDRAMでのスウィッチの働きと、イオンチャンネルの特性に応じた一種の抵抗が合わさった素子と考えることができる。
ポンプを使って電位を維持する限り、細胞は自分で発電し、細胞膜という電気を貯めるコンデンサとスウィッチとしてのチャンネルを持つことから、DRAMとよく似た構造をとっているのがわかる。神経細胞の場合、電位はチャンネルが閉じている限り電気維持されるが、チャンネルが開くと外界との勾配はどんどん消えていく。このように原理的には一個の神経が一つの記憶素子として働く可能性を持っているが、実際にDRAMのような使われ方をしているのかどうかについては私にもよくわからない。また、ポンプとチャンネルを備えている細胞は、神経細胞に限らず数多く存在する。したがって、このDRAMに似た構造だけが記憶に大事だとは言えないだろう。
神経細胞自体を記憶素子として使わずとも、神経細胞を組み合わせて回路を形成させ記憶させることも可能だ。エレクトロニクスでは様々な記憶回路がDRAMのような記憶素子の代わりに使われている。もっとも単純なものは図4に示すラッチ回路と呼ばれるもので、インバーター、ANDとORの演算素子を結合させた後、ANDからの出力に応じてフィードバックさせること回路で、0と1を記憶させることができる。
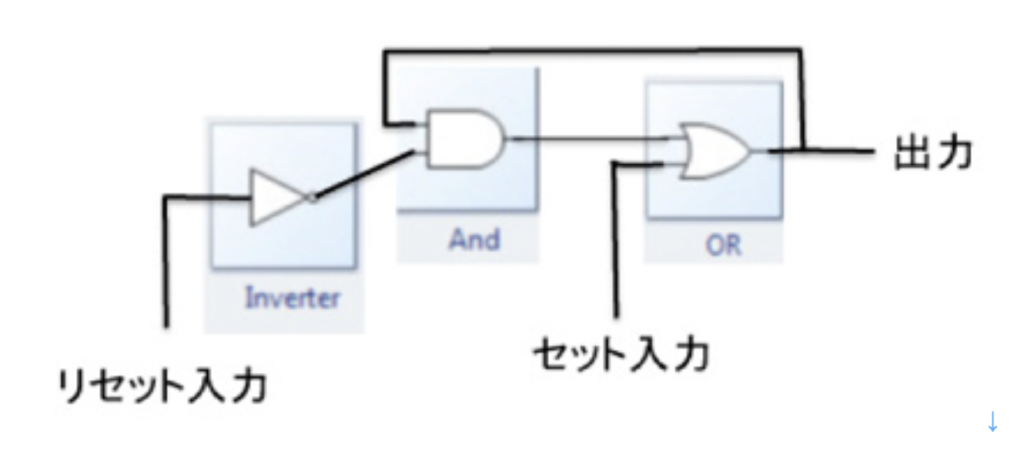
図4 最も単純なRSラッチ回路:説明は省略
電気回路の講義のつもりはないので、ANDやOR といった選択が行われる素子を繋ぎ合わせれば、それ自体で記憶が可能になると考えて貰えばいい。
しかし同じような回路形成を細胞で実現しようとすると、神経のように特殊な形態と機能を持つ細胞が必要になる。いくら興奮性があっても、筋肉細胞ではこのような回路形成は不可能だ。この回路形成が可能であるという点を念頭に神経細胞の特徴を述べたのが、細胞レベルでの脳研究の始祖、ラモンカハールのニューロン4原則だ。
カハールのニューロン4原則
- 1)第一原則:ニューロンは脳の基本構成成分であり、シグナル伝達単位である。
- 2)神経は軸索と樹状突起に存在する特殊な部分を通してのみ相互作用を行う。
(シナプスに似た構造をすでに考えていた) - 3)この特殊部位(シナプス)を介する一つの神経細胞の結合は、特定の神経細胞同士に限定されている。(結合の特異性)
- 4)シグナルは常に一方向のみに伝わる。(シグナルの方向性)
この原理は1890年に書かれたものだが、なぜ神経だけが、演算回路や記憶回路を形成できるのかの原理について見事に説明できている。このことは、図5に示したカハールの網膜神経組織の模式図からよく理解することができる。
(https://en.wikipedia.org/wiki/Santiago_Ram%C3%B3n_y_Cajal)
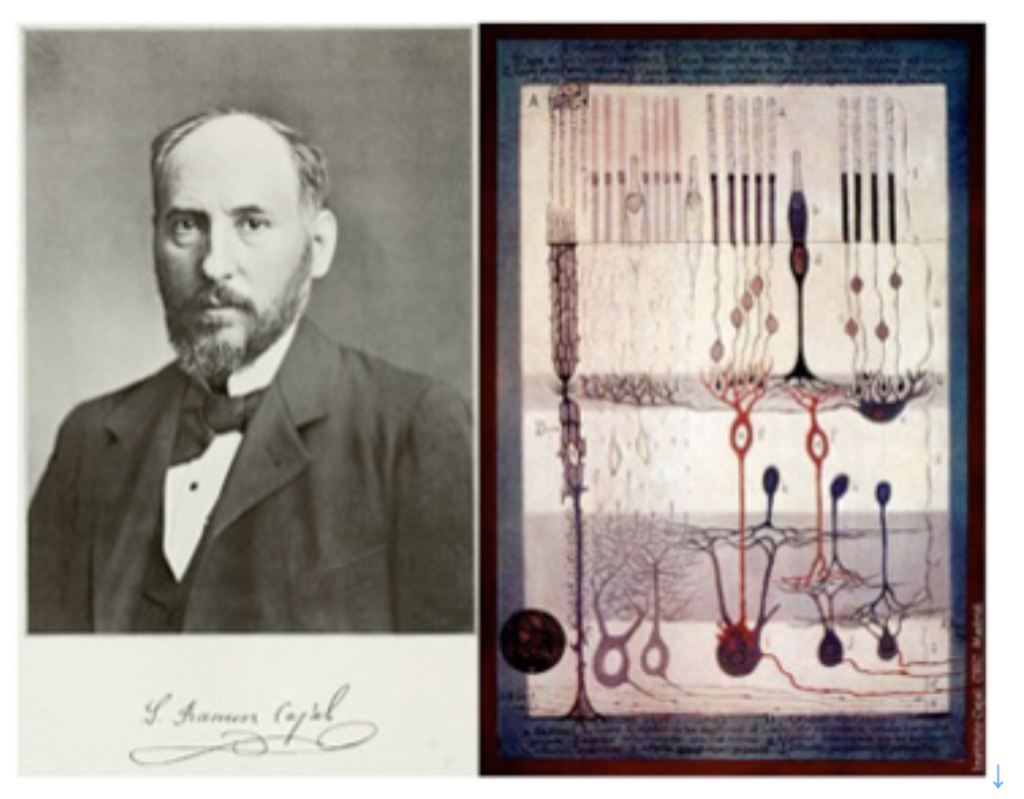
図5:カハールと網膜神経結合のスケッチ
すなわち、それぞれの神経細胞は、軸索や樹状突起の神経接合部のみで、しかも決まった神経細胞とだけ結合している。そして、感覚細胞からいくつもの神経を経て脳へとシグナルが一方向へ伝わっていることもよく描かれている。まさに、神経細胞は回路形成のために誕生した。
次は神経細胞が形成する記憶回路について、記憶の研究でノーベル賞に輝いたエリック・カンデルの研究を追いながら見てみよう。
[ 西川 伸一 ]
神経記憶III
2016年12月15日
神経科学の歴史について詳しいわけではないが、記憶を始めすべての脳機能が神経細胞(ニューロン)同士の結合した回路により担われていることを最初に示唆したのはスペインの生んだ巨人ラモン・カハールだろう。前回述べたように(前回図5)、カハールはゴルジ染色法を用いて、一個一個の神経細胞を染め出し、脳が様々な神経細胞のネットワークであることを明らかにし、カハールの4原則を提案した。このことから、カハールを神経細胞ドクトリン(ニューロン説)の最初の提唱者と呼んでいいだろう。実際この4原則に適合するネットワークを形成できる細胞は神経細胞以外にない。
カハールのニューロン4原則に加えて、神経回路を考える時に重要なドクトリンをあと二つ挙げておこう。
一つはすでに前回紹介したが、神経細胞は共通のメカニズムで、細胞膜上で起こる電気的興奮を伝搬することができる点だ。静止状態では、神経細胞内外のナトリウムイオン(Na+)勾配によって、神経細胞内外の電位差(静止膜電位)は-90mVに保たれている。外的な刺激でこの電位が変化してある閾値を超えると、イオンチャネル(電位に応じて開閉するNaチャネル)が急速に開き、結果急速にNa+が細胞内に流れ込み、膜電位の極性が変わる(+20mV:脱分極)。この電位の変化は、イオンチャンネルを介してK+を細胞外へ逃すことですぐに元に戻るが、刺激が繰り返されるとイオン勾配が消失して、興奮できなくなる(不応期)。このイオン勾配を復元するためには、それぞれのイオンに特異的なポンプが働く必要がある。このイオンドクトリンに従う細胞であれば、いかに複雑な形態をもち、神経間の興奮伝達に様々な化学物質が使われる場合も、神経を点から点に張り巡らせた電線と同じように単純化して考えることができるようになった。
最後のドクトリンが、神経細胞間の興奮伝達がシナプス接合部での化学物質の伝達によって誘導されるとするシナプスドクトリンだ。すなわち、化学物質の伝達によって、それを受け取る側の神経細胞の興奮を誘導できるが、この場合は電位依存性チャネル(voltage gated channel)ではなく、シナプスで興奮した細胞から分泌される伝達物質と結合することで開くチャネルにより、もう片方の細胞に興奮が伝わることがわかっている。シナプスドクトリンは、神経興奮伝達が一方向にのみ進むというカハールの原理を説明するだけでなく、使われる化学物質を変化させることで、神経間の結合に特異性を与え、また次の神経の興奮に対して促進的にも抑制的にも働けることが説明できる。この二つの性質を合わせると、ほとんど無限の神経回路を作れることが直感的にわかると思う。
この3つのドクトリンを基盤にして、神経回路による記憶維持の細胞学的、生化学的メカニズムについての研究が始まったが、この問題を分子レベルまで掘り下げて説明するのに成功したのがコロンビア大学のエリック・カンデルで、彼はこの研究によりノーベル医学生理学賞に輝いている。
カンデルが記憶回路の解明に用いた動物はアメフラシ(図1)で、大きな殻のないカタツムリと考えて貰えばいいだろう。

図1 アメフラシ(http://en.wiktionary.org/wiki/File:Aplysia_dactylomela_2.jpg)
カンデルは、図2に示した、アメフラシの水管(水を吐き出す管)を刺激すると、エラを引っ込めるという反射に注目した。図2は2000年カンデルがノーベル賞を受賞した時のプレスリリースに掲載されたアメフラシの図だが、腹側にエラと水管が存在している様子がわかる。カンデルたちは、エラを引っ込める反応をアウトプット、水管を触れる刺激、および尻尾への強い電気刺激をインプットとして、それぞれの刺激間の関係や、刺激のパターンが記憶されるかについて調べた。
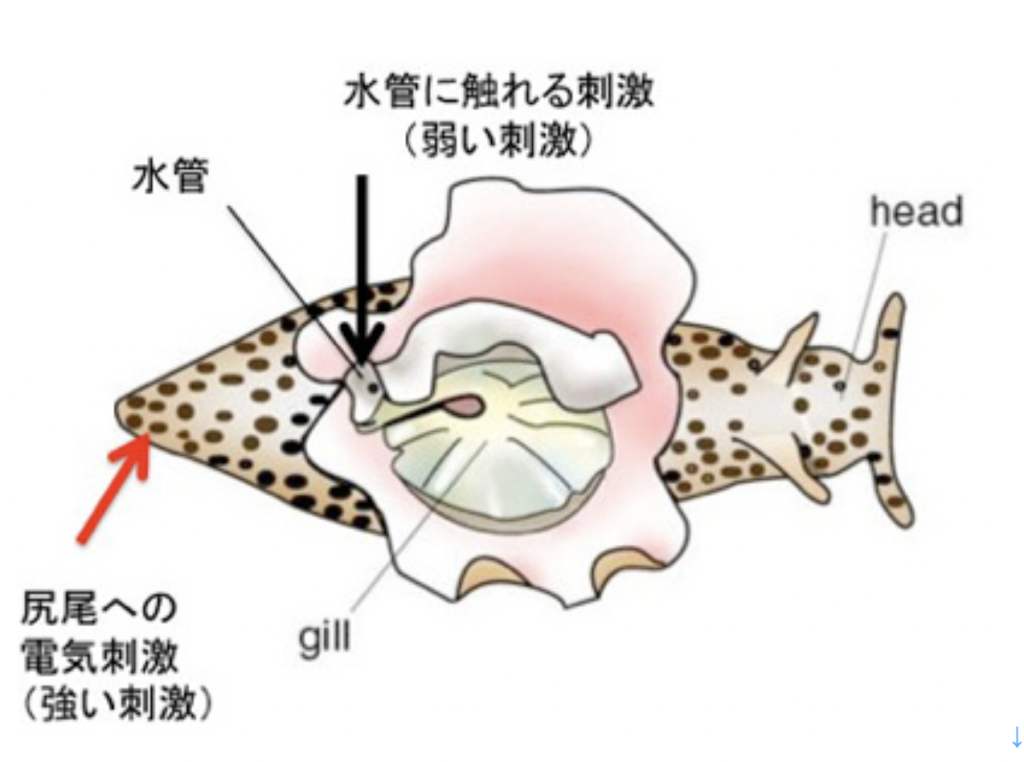
図2:カンデルの用いた実験系。(C) The Nobel Assembly at
Karolinska Institute
例えば水管を刺激し続けると、アメフラシは刺激に慣れてエラ反射を起こさなくなる(Habituation)。あるいは、水管に触れる前に尻尾に強い刺激を与えると、水管を触れた時のエラの反応が増幅する(Sensitization)。このサイフォン刺激や尻尾への強い刺激が記憶されているかどうかは、一定のインターバルをおいて、同じ刺激を加え、エラが同じ反応を示すかどうか調べることで確認出来る。このインターバルが短い時の記憶が短期記憶で、長い時の記憶が長期記憶になる。カンデルたちは特にSensitization(感作)と呼ばれる行動を用いて短期記憶、長期記憶について研究を行った。
もちろん短期記憶、長期記憶が存在することは、様々な神経システムで観察することができる。しかし、記憶に関わる神経回路を、一個一個の細胞レベルまで分離することは簡単でない。しかしアメフラシの回路では、神経が大きく、また数が少ないため、エラを引っ込める行動とともに、それに関わる単一の神経細胞の興奮を長期間記録し続けることができる。また、刺激後何日もたって、再度同じ神経を探し出して記録することも可能だ。記憶のメカニズムを、行動、神経生理学、そして細胞内での生化学的反応へと還元したいと考えていたカンデルにとっては、アメフラシは選びに選んだモデル動物だった。
図3はカンデルたちが分離した、水管反射の記憶に関わる神経回路の模式図だ。水管への刺激を感じる感覚神経と、エラを引っ込める運動神経がシナプス結合を形成しており、これがメイン回路になる。そこに、水管の刺激をフィードバックしたり、フィードフォワードする刺激性介在ニューロン、抑制性介在ニューロンが結合してサーキットが形成される。このサーキットに、尻尾の感覚神経と結合する調節介在ニューロンが結合して、体の他の部分の感覚を伝えるまさに回路が形成されている。
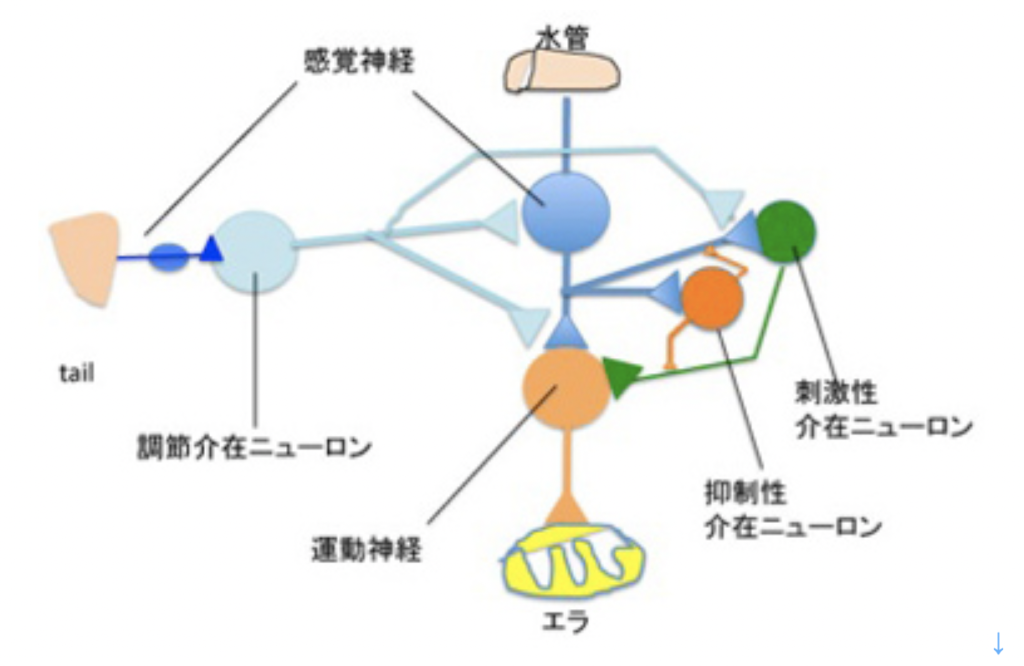
図3 カンデルたちが研究したエラを引っ込める行動を調節する神経回路。実際には、他にも調節介在ニューロンが関わっていることがわかっているが、ここでは省略する。基本的に、水管の刺激を感じる感覚神経と、エラを動かす運動神経の結合した経路に、抑制性および刺激性介在ニューロンが結合し、また全く異なる組織(tail)からの刺激が調節介在ニューロンを介して、この回路に伝えられる。
それぞれの神経は、単一細胞レベルで興奮を記録するとともに、物質を注入することができる。これにより、行動を神経生理学、そして最終的に神経内部での分子間相互作用へと還元することが可能になった。
カンデルたちの期待通り、現在ではSensitizationについての短期記憶、長期記憶のメカニズムはほぼ明らかになったと言える。両記憶のメカニズムについて、同じくノーベル財団のプレスリリースで使われた図を用いて説明しよう。
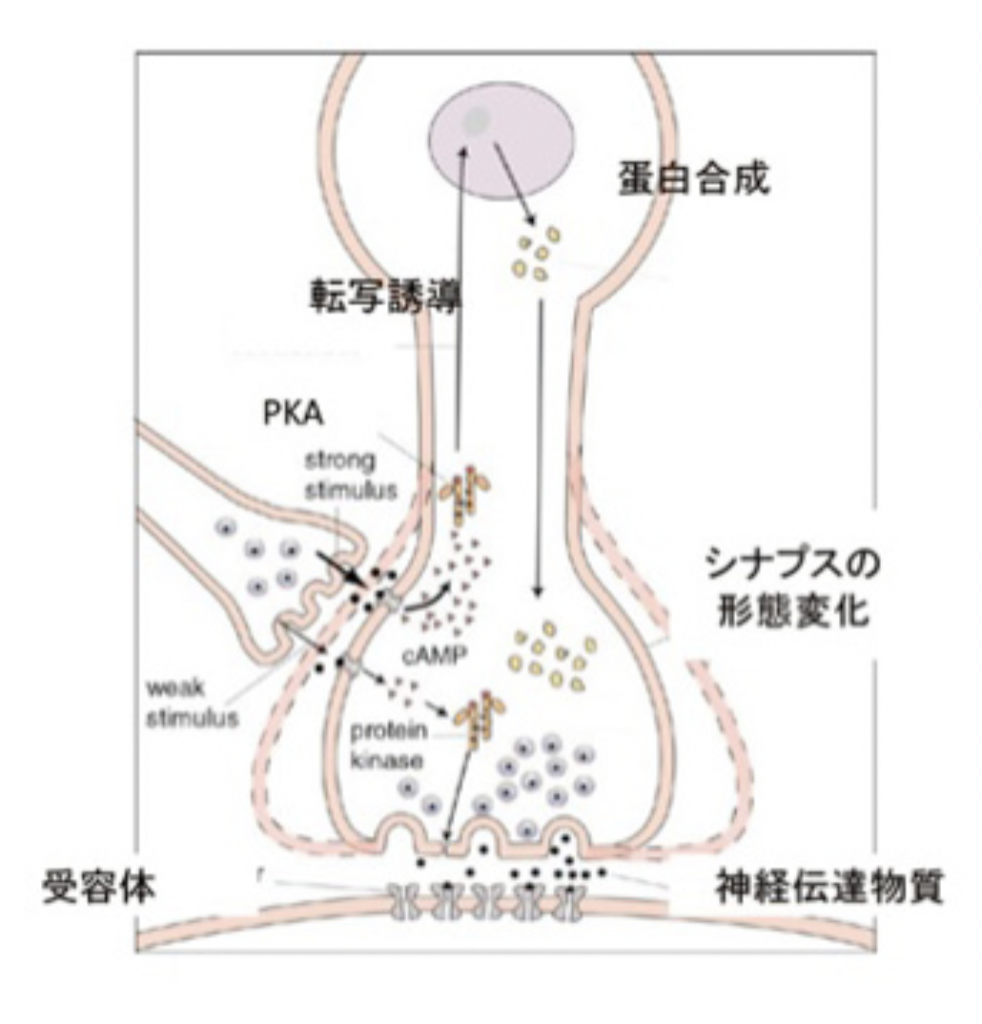
図4:短期記憶と長期記憶。(C) The Nobel Assembly at
Karolinska Institute:説明は本文
結論的には、「短期記憶も、長期記憶も、アウトプットとしてシナプス伝達の強さや閾値を変化させる細胞内の変化」とまとめることができる。
まず短期記憶から見てみよう。尻尾からの調節性介在ニューロンは、水管の刺激を感じる感覚神経と運動神経とを結合しているシナプスに結合している(図3)。この介在ニューロンが興奮すると、1)セロトニンを分泌し、2)このセロトニンがシナプス上の受容体に結合すると、3)アデニルシクラーゼが活性化し、4)cAMPが作られ、5)cAMPにより細胞内のタンパク質リン酸化酵素PKAが活性化され、6)カリウムチャンネルや様々なシグナル分子をリン酸化して、7)より多くの神経伝達物質を分泌するシナプスに変化する、という順序で運動神経への刺激が増強する。ただ、この増強は一過的なPKAリン酸化活性の増強によるもので、時間が経てば元に戻る。
これに対して長期記憶は、同じように活性化されたPKAが一過的にシナプス機能に関わる様々な分子を活性化するだけでなく、図4にあるように転写因子を活性化し、誘導された新しい分子により細胞の分化が誘導されることで、長期にわたるシナプスの大きさ数の変化が起こることで維持されることがわかった。実際には、セロトニン刺激がPKAを活性化するところまでは短期記憶と同じだが、長期記憶の場合はその後CREB1と呼ばれる転写因子が活性化され、新たな遺伝子を誘導することで、細胞自体が分化し、細胞及び組織レベルの構造変化が起こることで長く続く記憶が成立する。言い換えると、細胞をプログラムし直し、違った性質に変化させることが長期記憶のメカニズムだ。この仮説は、感覚神経へCREB1を過剰発現させることで、刺激なしでも長期記憶が成立するという実験による証明された。
実際にはこの単純な回路では、さらに複雑な短期、長期の細胞変化が起こり、シグナルのインプットとアウトプットの関係をさらに複雑にできることがすでに明らかにされているが、その詳細は省いていいだろう。
アメフラシの神経回路を用いた短期、長期記憶については以下のようにまとめることができる。
- 1)記憶は、複数の神経がシナプスで結合した神経回路を基盤に成立する。
- 2)シナプスから分泌される神経伝達因子は、細胞の興奮だけでなく、細胞内のシグナル伝達経路を刺激する。
- 3)このシグナルにより、一過性に細胞内の様々な分子が活性化、あるいは不活化され、その結果シナプス興奮の閾値や大きさが変化し、刺激に合わせた反応を誘導できる。これが短期記憶。
- 4)同じシグナルは、特定の転写因子を活性化し、新しい遺伝子発現を誘導する。これにより、細胞のプログラムが変化し、細胞分化や構造変化が起こる。一旦プログラムが書き換えられると容易に元に戻らないため、長期にシナプスの活性を変化させることができる。これが長期記憶。
以上のことから、長期記憶も、短期記憶も、特に特別なメカニズムを使っているわけではなく、多くの細胞と共通の分子を利用していることがわかる。PKAは多くの細胞でcAMP濃度上昇を感知し、短期・長期の細胞変化を誘導するのに働いている。CREB1についても同じだ。ただ神経の場合、刺激により誘導される最終結果が神経細胞同士の神経伝達の強度や閾値の変化になる。またこの変化も、一過性から、様々な長さの時間持続させることが可能だ。
次に、図3からわかるように、神経同士の結合はほぼ無限に複雑化することができる。しかも、神経細胞は形態的にも機能的にも何十種類も存在するため、回路の複雑性はすぐ天文学的になりうる。そして、神経伝達物質や受容体も数十種類存在するため、これによって可能なシナプスを介する興奮伝達の様式も無限に変化させることが可能になる。
しかしネットワークがどんなに複雑でも、背景にある生化学的原理はほぼ共通であることが重要で、どんな複雑な神経系回路でも、刺激が誘導するのは、回路自体の特性のリプログラムだ。
これまで、私の知識を再整理する意味で、神経生物学にあまりに深入りしたが、勉強はこのぐらいにして、次回は神経回路による情報と記憶の特性について考えてみたい。
[ 西川 伸一 ]
記憶と個性
2017年1月5日
ここまで、ゲノム、クロマチン情報、そして神経細胞ネットワークについて、情報と記憶という観点から検討してきた。今回はこれまでのまとめとして、3種類の情報システムを、個性という観点から整理してみようと思っている。
個性というと、「差異、多様性」だけを思い浮かべがちだが、例えばサルと人間の違いを個性とは言わない。個性というとき、あくまでも人間、あるいはサルの個体間で見られる違いについての話だ。したがって、個性という言葉の中には、種としての同一性と個体としての多様性の両方が含まれている。言い換えると、環境に適応するため種としての共通性を維持しようとエントロピーを低下させようとする方向と、ゲノム変異により個体を多様化しようとエントロピーを拡大する方向がぶつかり合うところに個性が成立している(図1)。

図1: ゲノム変異は常に種内のエントロピーを上昇させ、これに対し自然選択はエントロピーを低下させる方向に働く。この結果、種という制限の中で多様性が生まれ、個性が形成される。
おそらくダーウィンほど(ウォレスもだが)、この個性の理解が進化を理解する鍵であることがわかっている人はいなかっただろう。種という制限の中で、次の世代に伝えることが可能な個性が自然に生まれることを確信できないと、自然選択説はでてこない。
人間に個性があるように、あらゆる動物にも個性があることを一旦認めれば、産業革命下の弱肉強食社会を目の当たりにしていたダーウィン時代の都市のインテリにとって弱者が淘汰され強者が残るという考えは当たり前のことだったはずだ。一方、淘汰自体の概念と比べると、あらゆる生物が子孫に伝えることができる個性を自然に発生させるという考えは、背景にあるメカニズムが全くわからないため、受け入れることは簡単でなかったはずだ。もちろん当時の人も、人間を真面目に観察しておれば、それぞれが個性を持っており、誰一人として同じでないことはわかっていたはずだ。しかしダーウィン進化論にとって個性とは、子孫に伝わる性質でなければならなかった。メンデル遺伝学が登場するのは十年後だ。それでも、種としての共通性を基盤として、生殖過程で自然に発生する変異の蓄積が個性を生み、それが子孫に受け継がれると考えたダーウィンの慧眼には瞠目せざるをえない。
次にこの19世紀の議論を、21世紀ゲノム時代の視点で見直してみよう。
今や何百もの種の全ゲノムが解読されている。ゲノムの大きさ、DNA複製のメカニズムを知れば、大腸菌でさえ一回分裂して2個の大腸菌になるとき、両方のゲノムが全く同じである可能性があるなどと考える人はいないはずだ。すなわち、分裂時、あるいは分裂後に個体ごとに異なる様々な変異がゲノムにおこり、ゲノムは多様化し、個性が生まれる。変異は、1代、2代と世代を重ねることにより蓄積し、個体数の増加とともに多様性は増大する。このとき、増殖を損なう変異がおこると個体の維持ができずに自然に淘汰される。逆に、増殖力が高まって他の個体の個性を凌駕することも可能だ。このように、原核生物の場合、生存と複製に必要なゲノム構造が損なわれない限り、種内のゲノム多様性が発生し続け、こうして生まれた個性が環境にフィットすると、今度はこの個性が優勢になり、新しい個性が種として共通の性質になる。
ゲノムのみが経験(自然選択)を記憶できる媒体である原核生物では、ゲノムの多様化はエントロピーの増加を意味する。こうして増加したエントロピーは、自然選択過程により環境にフィットした個性が選択されることで低下する。この過程が異なる環境で起こると、選ばれる個性は異なるため、それぞれの個性へと種全体が収束することで、環境に応じた種が誕生し、種の数は増加する(もちろん環境が同一だと、種は分化できない)。このエントロピーが増減を繰り返す過程の繰り返しが進化で、すべてのゲノムは38億年前の生命誕生まで遡ることができる。このように、ゲノムに蓄えられた過去の記憶はゲノムの個性に等しい。言い換えると、個性とは過去の経験の記憶により形成される多様性と言っていい。(図2)
図2 個体のゲノムは自然に変異を蓄積するようにできている。これにより個性の多様化が起こる=エントロピーが増大する。環境に適応した個体が選択されると多様性は低下し、新しい性質が共通の性質になる。これが様々な環境で起こると、種は多様化する。
原核生物では、ゲノムの記憶がその生物の個性のすべてを決めていると言えるが、多細胞生物が生まれ、生殖細胞と体細胞が完全に分かれると、個体の個性は、ゲノムの個性とは言えなくなる。このことは私たち人間について考えるとよくわかる。
現在では同じ種でも個体間でゲノムが多様化していることがわり、確かにゲノムの個性が子孫に受け継がれ維持されていることは間違いがない。このゲノムの個性から、病気のリスクや、できれば他の様々な性質を予測したいと、個人遺伝子診断サービスが最近提供されている。しかし、個人の全ゲノムを解読したとしても、それですべての個性を予測できると考える人はいない。受精卵が発生を始めてから死ぬまで、我々は様々な経験を通して、ゲノムでは決まらない個性を作り上げていく。これは経験が記憶され、その記憶が個性となるからだが、この記憶はエピジェネティックスとして総称される。ただ、この時の記憶媒体は、クロマチン情報と神経ネットワークの両方(人間では言語も加わるが)からできている。例えば昔人気を博した一卵性双生児姉妹、金さん銀さんの性格や顔があれほど異なるのは、クロマチン情報の記憶と神経ネットワークの記憶の違いが原因だ。
まずクロマチン情報を介する個性について見てみよう。ゲノムの個性と比べた時クロマチン情報を介する個性を研究する難しさは、個体レベルの個性が一つの情報に代表されるのではなく、細胞一個一個の個性の集合として表現されている点だ。
まず発生過程で、何百種類もの細胞が分化するのは細胞ごとにクロマチン情報の違いが生まれるからだ。ただ、発生過程では、決まった細胞を繰り返し安定に分化させることが要求され、クロマチン情報が間違って多様化しないように設計されている。もちろん、この過程でも間違いは起こり、その間違いが細胞の生存や機能を損なわない限り、細胞の記憶として維持される。以前述べたように、クロマチン情報も細胞分裂を超えて維持することができるため、発生の初期に起こった間違いは、多くの子孫細胞に受け継がれる。このような間違いは、時間を経て問題になる場合がある。
有名な例は、1944年の冬、ナチスドイツに封鎖され極度の飢えに苦しんだアムステルダムで胎児期に飢えを経験して生まれた人たちは、高齢になってからインシュリンの分泌が低下し糖尿病になる。これは、胎児発生時に飢えにさらされて起こったクロマチン情報の間違いが、高齢になってから細胞に影響し始めたことを示唆している。同じように、胎児期にお母さんがアルコールを摂取すると、様々な細胞でクロマチン情報の記憶間違いが起こることが知られている。
幸い、クロマチン情報の記憶形成は間違いが少ないように設計されており(増殖因子や分化因子がきめ細かく細胞の方向性を調節する)、それぞれの細胞でおこる記憶間違いはランダムに起こるが細胞の生存から見て許容範囲内で止まっている。またクロマチン情報の変異は細胞ごとの個性になるため、変異が許容範囲を超える細胞は淘汰されるが、個体全体には影響が少ないことが多い。
同じ臓器で同じ細胞が作り続けられるためにはクロマチン情報は必須で、それを維持することは重要だが、発生後も外界のストレスによりクロマチン情報は少しづつ変化する場合が多い。例えば、老化によりクロマチン情報が変化することが知られている。しかし、このような変化は個々の細胞レベルで止まり、各細胞で全くランダムに起こるため、臓器や個体の機能にとって問題にならない。しかし、この個々の細胞にランダムに蓄積した様々なクロマチン情報の変化が集合したものが、クロマチン情報を介する個性として表現される。
以上をまとめると、クロマチン情報の記憶は、各細胞個別に形成される。この記憶は細胞の種類に応じて同一になるよう設計されているが、どうしても小さな記憶違いが生じて、これが細胞の個性を形成する。そして、この細胞の個性が集まって、個体の個性になるが、ゲノムの個性と異なり、個体レベルのクロマチン情報の個性を一つの情報で代表させることはできない。また、クロマチン情報を介する記憶や個性は、各細胞に限局されており、統合されることはない。個体レベルのクロマチン情報の個性は、あくまでも細胞の個性がただ集まっただけの結果だ。
エピジェネティックスな変化の中には、神経ネットワークを介する情報の記憶も当然含まれる。事実、私たち人間の個性は、脳の個性として表現される部分が多い。例えば、生後訓練を繰り返して筋肉が発達し、早く走れるようになる過程にアメフラシで見たのと同じ神経ネットワークを介する記憶が大きく寄与している。もちろん、筋肉自体も発達し、これにはクロマチン情報の変化も寄与するが、神経系の記憶が存在しないと筋肉は発達しない。残念ながら、こうして訓練した結果は、次の世代に受け継がれず、一代限りだ。
神経ネットワークのメカニズムを突き詰めると、細胞が発生し、成長し、老化する過程で起こるクロマチン情報の記憶と同じと言っていい。どんなに複雑な脳神経系でも、進化過程でゲノムに蓄積された記憶情報に基づいて空間的・時間的に発現する特異的なシグナルに従って順々にクロマチン情報が変化した細胞を分化させ、これをゲノムの記憶に従って立体的に構造化させることで、形成される。また、生前、あるいは生後それぞれの神経が受ける刺激に対しては、一次的な反応が細胞内シグナル伝達経路の活動として反応する。そしてシグナルの質や強さに応じて、前回見たように、神経細胞のクロマチン情報が変化し、長期記憶が成立する。しかし図3に示すように、神経細胞の反応は他の体細胞の反応と全く変わるところはない(図3)。
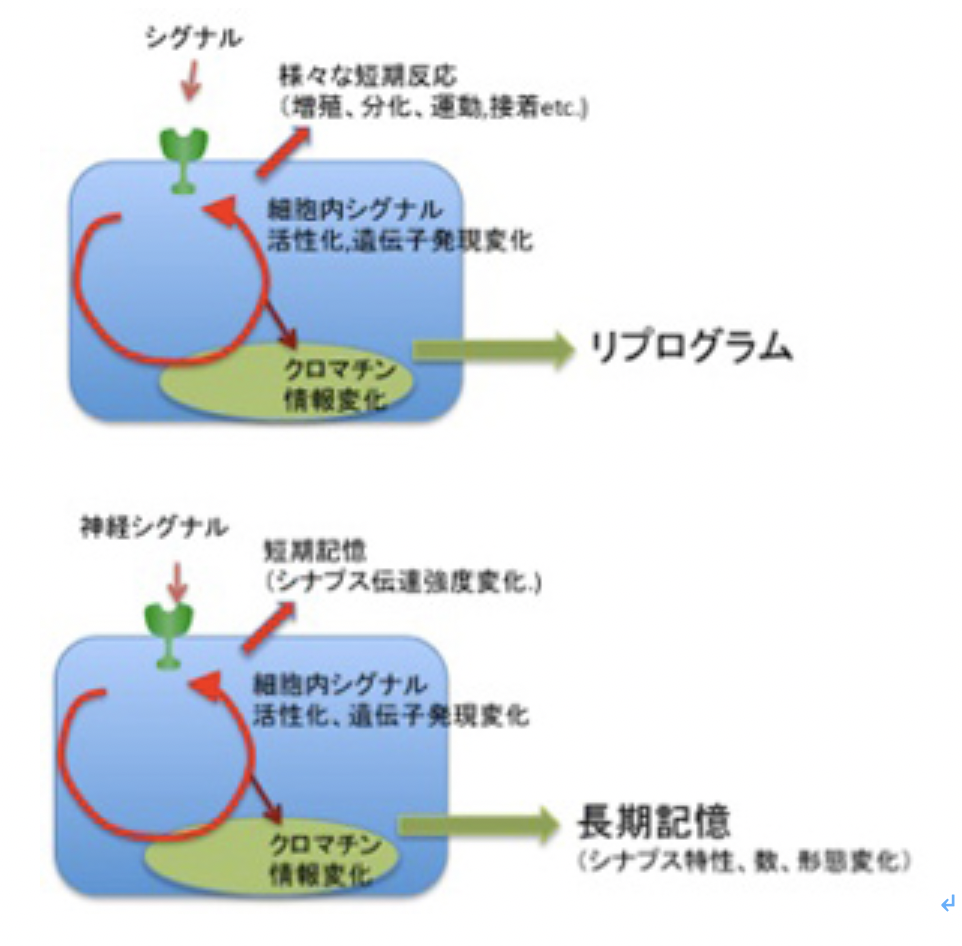
図3:神経細胞と一般体細胞の比較。
説明は文中。
一方、一般の体細胞と神経細胞の記憶は、個々の細胞内で起こる変化が個体レベルで統合されるかどうかという一点で大きく異なっている。すでに述べたように、クロマチン情報の記憶に起因する細胞の個性は、ただ集合して個体の個性に寄与するだけだが、神経系では細胞レベルのクロマチン情報の変化に起因する個性は、細胞同士がネットワークを形成することで、個体レベルで統合される。この統合は、神経細胞にとどまらず、筋肉やホルモン産生細胞など、多くの体細胞をも統合できる。
実際には、同じような細胞の変化を統合して個体の個性を変化させることは、内分泌システムにも見ることができる。オタマジャクシがカエルへと変態を遂げる時、甲状腺ホルモンが分泌され、体全体の細胞を統合する。同じことは、私たちが思春期に急速に様々な性的特徴を発生させるのも同じだ。しかし、一見個体全体を統合している高等動物の内分泌システムを詳しく見てみると、ほとんどがどこかで神経系とつながっており、神経ネットワークの記憶の支配を受けていることに気づく。
このように、神経細胞は、神経細胞にとどまらず様々な細胞と直接、間接にネットワークを形成する能力を開発し、全く新しいレベルの個体性の統合を可能にしている。
これに加えて、神経ネットワークでは、視覚、聴覚、嗅覚など、外界の変化を迅速に、立体的に捉え、この入力を個体として統合された神経ネットワークにリンクさせることができる。これにより、環境を全く異なるレベルで自己化することが可能になった(例えば景色の記憶は、環境を自己化したと言える)。もちろん、外界の自己化は原始ゲノム発生以来、生物進化の駆動力と働いてきたことだが、神経系による自己化は、あるがままの外界を表象し自己の延長として自己化する点で、これまでの自己化とは全く異なる。
この個体内外の統合と、自己の発生についてはわかりにくいと思うので、次回は、「統合と自己」というタイトルで、神経系ネットワークが可能にする個体と細胞、個体と外界の統合能力について解説してみたいと思っている。
[ 西川 伸一 ]
統合と自己:細胞の個性と個体の個性
2017年1月16日
今回は個体と細胞の関係を考えながら、神経情報の特殊性について見てみる。
教科書的には、多細胞体制の誕生は細胞の個性と個体の個性の分離をうながし、またその結果個体のゲノムを継承するために生殖細胞が誕生したことになる。しかし、生殖細胞が誕生した後も、有性生殖だけが個体のゲノムを継承する方法ではない。例えばプラナリアのように個体全体が分裂する場合もある。植物では体細胞からカルスが形成され、そこから完全な植物個体を形成することができる。

図1:プラナリアは成長すると個体自体が分裂することが古くから知られている。(出典 https://www.flickr.com/photos/internetarchivebookimages/20705306246/sizes/o/)
有性生殖と無性生殖が共存できるのは、ほぼ同じゲノム情報が全ての細胞に存在しているからに他ならない。もちろん、細胞ごとにゲノム情報の小さな違いは生じているが、原則的にほぼ同じゲノム情報が個体の全ての細胞に存在していると考えていい。無性生殖を行わない動物でも、体細胞の核移植によるクローン動物作成実験から(哺乳動物のように複雑な動物でも、体細胞のゲノムを未受精卵に移植すると、新たな個体が発生する)、全ての細胞に完全なゲノム情報が存在していることが示された。この意味で、ゲノム情報の同一性が、細胞と個体の統一性を保証していると言える。
しかし、植物やプラナリアでは体細胞から個体を形成できるのに、私たち人間の細胞は、決して個体を形成することはない。この理由の一つは、私たち人間の細胞は、ゲノムは受精卵と同じでも、体細胞への分化過程で変化した細胞それぞれのクロマチン情報を簡単にはプログラムし直すことが難しいことだ。実際プラナリアも同じで、分化した細胞は他の細胞に分化し直すわけではなく、全細胞のうち3割がクロマチン情報を変化させやすい多能性幹細胞でできているからだ。同じゲノムを持っていても、不可逆的にそれぞれのクロマチン情報が選び取られて発生が進むことを、英国の発生学者ウォディントン(C.H. Waddington)はエピジェネティック・ランドスケープ(epigenetic landscape、後成的風景)という絵を用いて表現している。しかし、分化細胞を他の種類の細胞へと変化させることが難しいお陰で、私たちは明日皮膚が血液に変わってしまっているという心配をしなくて済む。
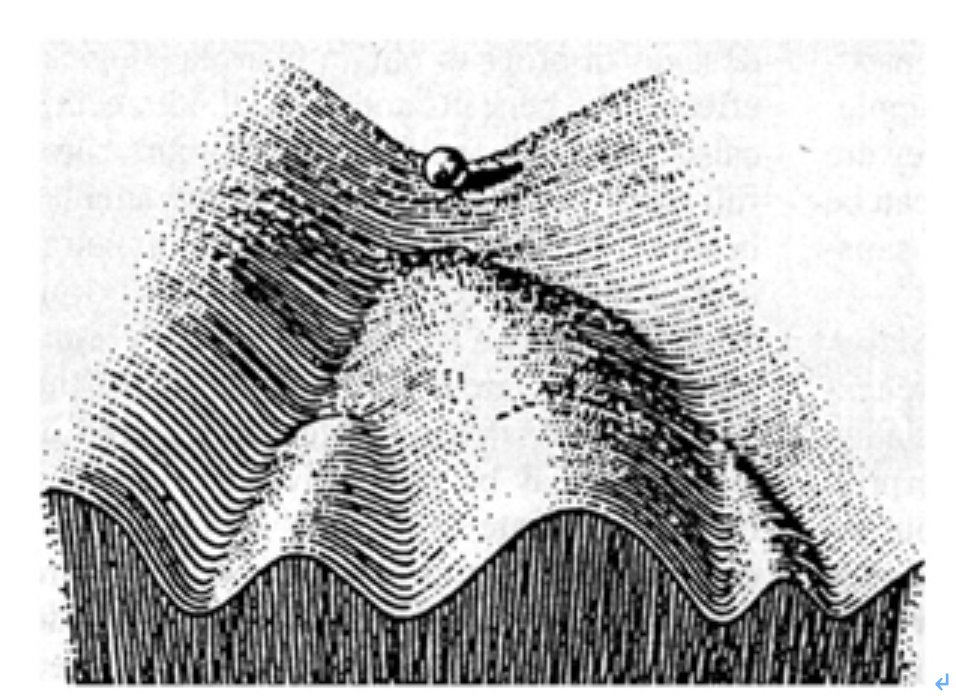
図1 ウォディントンの描かせたepigenetic landscapeの絵をわかりやすく書き直した図で、この概念を示す際、最もよく使われる。上部にあるビーズは様々な場所に動き得るが、一旦分化の方向性が決まると、他の道は選べないことが示されている。
結局、我々人間では、受精卵から始まる一生で一回きりの、ゲノムの指示に従う発生過程を通す以外に、必要な細胞を準備することはできない。ES細胞やiPS細胞の研究から、受精卵に限らず、体のあらゆる細胞を作る能力を持つ細胞が得られることが明らかになり、他の細胞から区別して多能性幹細胞と呼ぶようになっている。しかしあらゆる細胞に分化できる多能性幹細胞の塊から始めても、正常な個体を発生させることはできない。なぜなら、動物の発生過程では、個々の細胞が個体との関係を読み取り、これにより得られる情報の指示に従ってクロマチン情報を変化させることが要求される。例えば四肢はほとんど同じようなメカニズムで体幹から飛び出すが、頭に近いところにある細胞から腕が、遠いところにある細胞から足ができる。これも、各細胞がボディープランから見て最もふさわしい組織の形成を指示する、すなわち適切なクロマチン情報を確立するための情報が体の各部に用意されているからだ。しかし、体の体制を構成しているのも細胞だ。どちらが原因で結果かがわからないという因果性の問題が発生学には常に存在している。
ゲノムには、発生に応じて必要な遺伝子セットを発現させるためのプログラムが存在し、多能性の幹細胞は発生に合わせて様々なクロマチン情報記憶を展開することができる。多能性幹細胞は様々な条件下でほとんどの細胞へと分化できるが、秩序ある個体を形成するためには細胞の多能性だけでは不十分で、細胞が初期胚が持つ個体の体制からのシグナルを受けることが必要になる。例えばES細胞を集めて個体に移植したり、あるいは試験管で培養したりしても、奇形腫と呼ばれる秩序のない構造ができるだけで、マウスはできない。しかし同じES細胞を胚盤胞に注入すると、胚盤胞の体制に組み込まれてES細胞は正常個体を形成することができる。
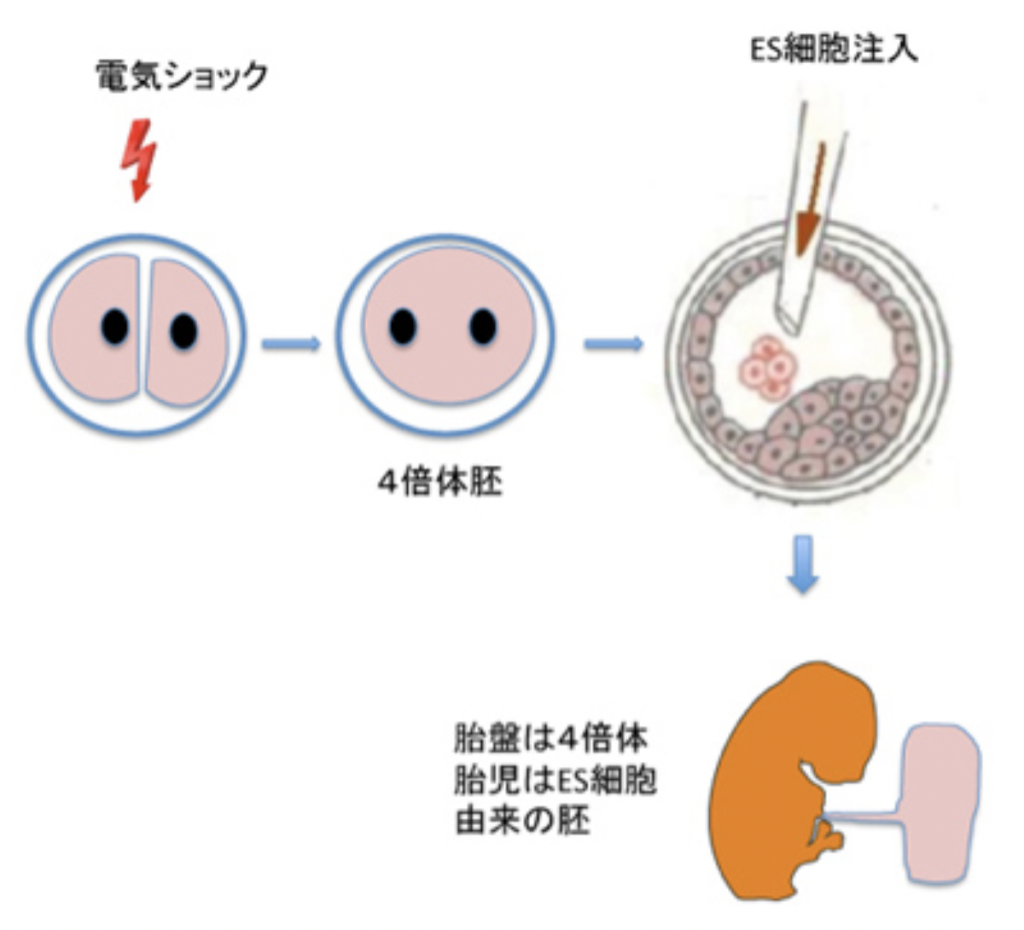
図3 4倍体補完法の説明 説明はテキスト参考
胚盤胞の体制自体が持つトップダウンの力を最もよく示す例が4倍体補完法と呼ばれる方法だ(図3)。受精卵が分裂した時に電気ショックで細胞を融合させると4倍体の胚ができるが、この胚盤胞をES細胞の塊に指示を与える構造として用いることができる。4倍体の胚は胚盤胞期まで発生するが、胎児細胞はそれ以上発生ができない。一方胚外のトロフォブラストは機能的胎盤を形成できる。このため胚盤胞に移入したES細胞やiPS細胞は、死滅する4倍体の細胞の代わりに発生する。すなわち、胎盤は4倍体の細胞から、胎児部分は注入したES細胞からできた合成胚を作り、発生させることができる。単純な構造に見える胚盤胞にも、細胞の分化に指示する体制が存在することがわかると思う。
このように発生学は、細胞の分化を指示する個体の体制とは何か?そのシグナルを受けた分化細胞がどのようにして個体へと統合されるのか?など、常に全体と部分の関係を研究してきた。このことは、例えば古典的な発生学で、オーガナイザー(organizer)とか、誘導物質(inducer)、ボディープラン(body plan)という個体の体制を示す言葉が重視されていることをみれば理解できる。この発生学の特殊用語を使ってまとめると、ゲノムの記憶を正確に展開するためにも、最初はオーガナイザーが必要で、このオーガナイザーもまた細胞からできており、ゲノムに指示により形成されるのだが、卵の形成過程、受精過程、着床過程など外部の要因が関わって形成される。
このように「卵が先か、鶏が先か?」の問題と同じで、発生では全体と部分は一体化しており、原因と結果の区別を困難にする。同じように、発生でのゲノム情報とクロマチン情報の関係も一体化している。発生でのクロマチン情報の変化は、ゲノムにコードされたプログラムで進むが、クロマチン情報がないと複雑な多細胞生物のゲノムの記憶を展開することができない。
では前回述べた多細胞動物に見られるゲノム情報、クロマチン情報の個性とはなんだろう。発生過程で一つの系列内の細胞の多様性が生まれることは、ゲノムやクロマチン情報の間違いと考えていい。これらの情報の変異は細胞ごとに起こるが、許容範囲以上の間違いは致命的になるか、細胞レベルで除去される(例外と言える免疫系は後で説明する)。発生、成長の過程で個々の細胞レベルで発生する個性は、個体の個性として統合されることはないが、細胞レベルの個性が個体に影響を及ぼす時、病気、老化などの異常として現れる。
少し長く説明しすぎたが、神経情報が誕生するまでは、新しく生まれた情報システムも、それ以前に存在した情報から独立することができていなかったことを示すためだ。これに対し、新しく生まれた神経情報は、他の情報に依存した生命活動から、大幅な独立を勝ち取っている。
もちろん神経ネットワークも、その形成や維持はゲノム情報、クロマチン情報に完全に依存しており、神経情報だけが独立することはない。体がないと、脳は機能せず、神経情報維持できず、心もできない。しかし、神経情報のオペレーションを詳しく見てみると、それ自体で他の情報から独立して働くことができ、また各細胞に生じた個性がそのまま神経系全体の個性として統合されていることがわかる。
例えば、一卵性双生児間の個性を考えてみると、細胞レベルのクロマチン情報の違いが集まって生まれる個性の違いと比べ、神経ネットワーク情報に起因する性格や記憶の違いは際立って大きい。すなわち、神経ネットワークの枠組みが一旦形成されると、神経情報のオペレーションは自由度が高く、結果多様な個性を形成できることを意味している。個性が多様化できるということは、他の情報からの独立性が高いことだ。この独立性が、デカルト以来私たちが心身二元論に陥る理由の一つだと思う。
しかしなぜ身体の死とともに消滅する運命の神経ネットワークの情報が、心身2元論という錯覚を生むほどの独立性を獲得できたのか?その原因について最後に考えてみよう。(以下は私の妄想でしかないことを断っておく。)
神経ネットワークが持つ最も重要な特徴は、様々な神経細胞があるにもかかわらず、各神経細胞の興奮、細胞内(軸索や樹状突起)での興奮の伝搬、そして神経間の興奮伝達のための原理がほぼすべての神経細胞で共有されている点だ。そのおかげで、視覚であれ、嗅覚であれ、触覚であれ、全てのインプットを同じ神経ネットワークで共有することができる。また、インプットに対するアウトプットも、筋肉を動かす運動からホルモンの分泌に至るまで実に多様で、多くの組織を直接間接に支配してネットワークの中に組み込むことができる。もちろん、インプット・アウトプットだけでなく、神経同士が刺激・抑制し合う回路も形成される。同じ原理で作動できるため、神経細胞の数さえ増やせば、無限に神経細胞同士を組み合わせ複雑なネットワークを形成することができる。こうしてできた神経ネットワークも、神経間の結合の特異性や強さを変化させるためにはゲノムやクロマチン情報に依存したメカニズムを用いるが、クロマチン情報が「ゲノムのon/off」のための情報であるのとは全く異なり、形成される記憶自体は、直接ゲノムやクロマチン情報と関係はない。
このようにゲノムとクロマチン情報に依存して発生が進み一旦回路が形成されると、あとは外来の刺激が回路のパターンを変化させる例は、すでにアメフラシの水管反応で説明した。アメフラシ水管反応回路では、刺激を受けなかった回路と、刺激を受けた回路では全く回路特性が異なる。これは回路構造の中の神経細胞の特性が変化したためだが、この変化は回路全体の個性として統合される。このように、神経回路はこれまでの情報と比べて、高い自立性を持ち、興奮伝達の原理を共有することで無限に複雑化する可能性を獲得した。しかも、さらにこの共通性のおかげで、個々の細胞レベルで生じた神経細胞の個性はネットワークにより共有されることで、神経系全体の個性へと統合される。
細胞の個性を統合することができる神経系の誕生は、刺激が消えた後もその刺激の影響を維持することを可能にした。この点については、同じようにほぼ無限と言える外来抗原に反応できる免疫系と比べてみるとよくわかる。
免疫系は無限の化合物に対して反応できる。これは、多様な抗原の一つ一つに対して反応する個別のリンパ球が体内に存在するからだ。すなわち、同じ細胞を繰り返し作り、細胞に個性が生まれるのを極力拝する他の組織と異なり、リンパ球は最初から異なる個性(抗原特異性)を持つよう分化する。この個性はゲノムレベルの個性で、抗体やT細胞受容体遺伝子を不可逆的に変化させ、それぞれのリンパ球に異なる抗原受容体遺伝子を発現させるリンパ球特異的な遺伝子再構成メカニズムに依存している。しかし個々の細胞レベルの免疫反応は抗原に反応する細胞だけでとどまる。すなわちリンパ球同士の反応には必ず抗原の介在が必要で、抗原がなくなると細胞間の相互作用は消滅する、一方、神経の場合は刺激が消えても、細胞相互作用は維持できる。
ニールス・イェルネらは、抗体が他の抗体により認識できる(イディオタイプ)という現象を元に、リンパ球同士は抗原を介さずに、神経のように直接相互作用できるとするイディオタイプネットワーク説を唱えた(図4)。しかし、抗体が認識し合ってネットワークを作るという考えはは最近ではあまり支持されなくなっている。結局、免疫系が無限の抗原に反応できるのは、ネットワークとは無関係に、リンパ球の抗原反応性に最初から多様性があるからだ。結局免疫反応とは、抗原反応性の異なる個別の細胞ごとに増殖、分化寿命延長などの細胞学的変化が起こることに他ならない。
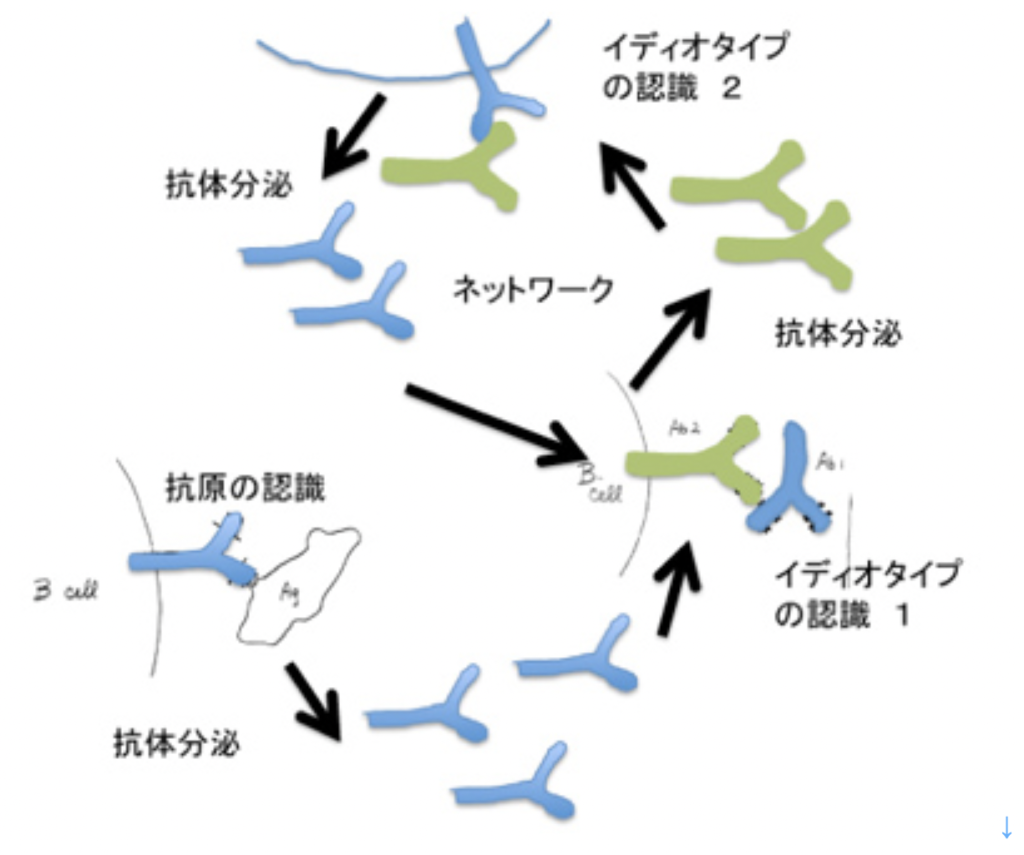
図4 ニールス・イェルネらが考えたイディオタイプネットワーク説。彼のノーベル賞受賞論文を元に書き直した。最初抗原により刺激されたB細胞から分泌された抗体1(空色)の抗原結合部は(他の抗体分子とアミノ酸配列が異なるため)抗原としても働き、抗体2(緑色)により認識される。この相互刺激関係が成立すると、抗原なしに刺激は持続する。
一方、イェルネが考えたように、免疫系でネットワークが成立するためには、最初の刺激が消えた後も、リンパ球同士で刺激しあえるメカニズムが必要だ。免疫系では、図4に示したことは理論的には可能でも、現実には特殊な状況だけで起こっているようだ。
一方図4に示したネットワークは神経系では成立できる。神経系も、味覚や嗅覚として化学物質を感じ、記憶できる。化学物質に反応する点では、免疫系と同じだが、例えばワインを一口飲んで、実際の刺激は胃の中に消えてしまっても、味や匂いを他の感覚と関連させ、記憶として維持することができる。これは、刺激が消えた後も、匂いや味の刺激によって生まれた新しい回路の活性が維持できることを意味している。これが可能なのも、神経細胞が同じ原理を共有し、回路を形成できるからだ。
以上、全ての神経細胞が興奮のための原理を共有することで、神経細胞同士で無限に複雑なネットワークを作り、また各神経細胞の変化を、ネットワーク全体の変化として共有できるようなったことで、心が身体から離れてしまうという心身2元論の幻想が生まれるほど、神経ネットワークによる情報が、ゲノムやクロマチン情報から独立した、独自の情報系を作ることができたことを納得していただいただろうか。
次回からは、各細胞の変化が統合可能で、他の情報から独立している神経情報系により可能になった脳新しい情報機能について見ていきたい。
[ 西川 伸一 ]

