
1、ゲノム科学
5、言語の誕生
6、文字の歴史
カテゴリ:生命科学の現在

2018年4月16日
今回から文字の誕生とその歴史を考える。ただ、ここで文字と表現しているのは、Writing(書くこと)という大きな意味で使っていく。
現在世界には6000を越す言語があるが、では文字はどのぐらいあるのかと考えると、世界中探しても、現在使用されている文字体系は数えられる程度しかないように思う(一応学校で教えている体系だった文字のみを想定している)。漢字とともにどの国でも使っていない表意文字を持つ日本や韓国に暮らしていると、言語一つ一つに異なる文字があるように思ってしまうが、例えばヨーロッパ、北米、南米、オーストラリア、アフリカ諸国を見てみると、キリル文字も含めアルファベットのバリエーションとして片付けることができる。詳しく見れば各言語に対応する文字にも少しずつ違いはあるが、それでもそれぞれを歴史的に遡ればほぼ同じ起源に行き当たる。もちろん、アフリカやアメリカのように、現在は使われていないが、独自の文字を持っていた言語も多い。ただ、このような独特の文字体系ですら起源をたどると、メソポタミアに残された文字まで辿れるのではないかと言われている。このようにそれぞれ独立に発生したと考えられる多くの話し言葉(ここではS言語と呼ぶ)と異なり、文字は生物の進化と同じで、階層的系統樹を持っている。
なぜ同じ言語の媒体なのに、これほどの差があるのだろう?答は以下のように言えるだろう。
「S言語と文字は独立して発生しそれぞれの言語の中で統合され新しい言語体系を形成している。しかし、一人一人の個人にとっては常にS言語が優先されてしまう構造になっているからだ。」
すなわち、私たちが生後最初に触れるのがS言語で、視覚による文字が文字として認識されるのはずっと後だ。さらに誤解を恐れず単純化すると、文字はどんなS言語にもなんとか合わせて使うことができる。
このことは我々日本語を考えればすぐわかる。日本語には全く独自の表音文字が2種類も存在する。しかし、これがなくても、また中国から受け継いだ漢字がなくても、ローマ字を日本語の文字として利用することは明日からでも可能だ。実際、明治政府発足後、当時の文部大臣森有礼がこの可能性を真剣に考えたと聞く。ひょっとしたら、これから漢字を習わなければならない小学生なら、全部ローマ字に変えることを断固支持するかもしれない。しかし、同じ小学生に、明日から英語にS言語を換えると言ったら大反対するだろう。このように、例えばローマ字は音を表す道具として、少し改良を加えればどの言語にも対応できるよう発達してきている。また、文字を覚え出すのは早くても3歳以降だ。一方、S言語の大枠は3歳までに頭の中に形成されている。このため、文字は常にS言語に対して従属的関係になる。
しかし一旦文字を使い出すと、文字は大量の情報を扱うことを可能にする。実際、文字を習うために私たちは脳のかなりの部分を犠牲にすることが、失読症の研究からわかっている。この文字により扱える情報量のおかげで、現在私たちが使っている言語体系のかなりの部分は、文字を介して形成されて来た。このため、おそらく我々成人の脳内に存在する言語体系でも文字の地位は、S言語をはるかに凌駕するようになっているのと思う。このように、言語はS言語、あるいは文字だけで定義できるものではなく、実際にはジェスチャーも総合された体系として見るのが正しい。
以上のことから、
などが、S言語と文字の関係として見えてくる。このことから、文字の歴史を知ることは、S言語とは独立して発達してきた言語(コミュニケーション体系)が、S言語と一体化して新しい言語体系を誕生させる過程を知ることと同じであることがわかる。
先に述べたように、様々な遺物から文字の起源をたどると、メソポタミアの絵文字にまでさかのぼれることが知られている(次回以降見ていく)。最初は意味を持つ文字(logogram)としてスタートしても、漢字を除いてほぼ全ての文字は、表意文字を捨て音やシラブルを表す文字へと転換を遂げる。これを概観するのが今回からのテーマだが、この歴史を学んでみると、現在我々が使っている日本語は、表意文字が表音文字へと変化していった多くの文字の歴史をそのまま残している、いわば生きた化石のように思えてくる。そこで、文字の最初は日本語を見直すところから始めたい。この生きた化石のような文字を使っている珍しい民族だからこそ、日本人は、表音文字(phonogram)のみを使っている人たちより、文字の成立過程や、S言語と文字の関係を、はるかに深く理解できるのではないかと思う。
まず、文字がS言語に固有のものではないこと、すなわち独立して誕生していいことは、日本語にとって文字が中国大陸からの輸入品であることからわかる。文字は、S言語に固有のものである必要はない。しかし、独立して発生した文字とS言語は、互いに統合された一体化した体系になる必要がある。日本語で言えば、中国の漢字に加えて平仮名とカタカタを誕生させ、漢字も日本語に固有の文字に生まれ変わらせたことがこれに相当する。同じように、ベトナム語のチュノムも、漢語をベトナムのS言語に合わせるために形成された文字だ。ただ、チュノムは漢字を単純化することなく、ベトナム風にアレンジして使ったため、大衆化が難しかった。そのためフランスによる占領時代に、結局チュノムや漢字は排除され、現在ではアルファベットをベースにした独自の文字が出来上がっている(図1)。
このように、文字とS言語は独立して誕生し、S言語に合わせて文字が変化することが日本語最初の文字が漢字だった、韓国語やベトナム語を見るとよくわかる。
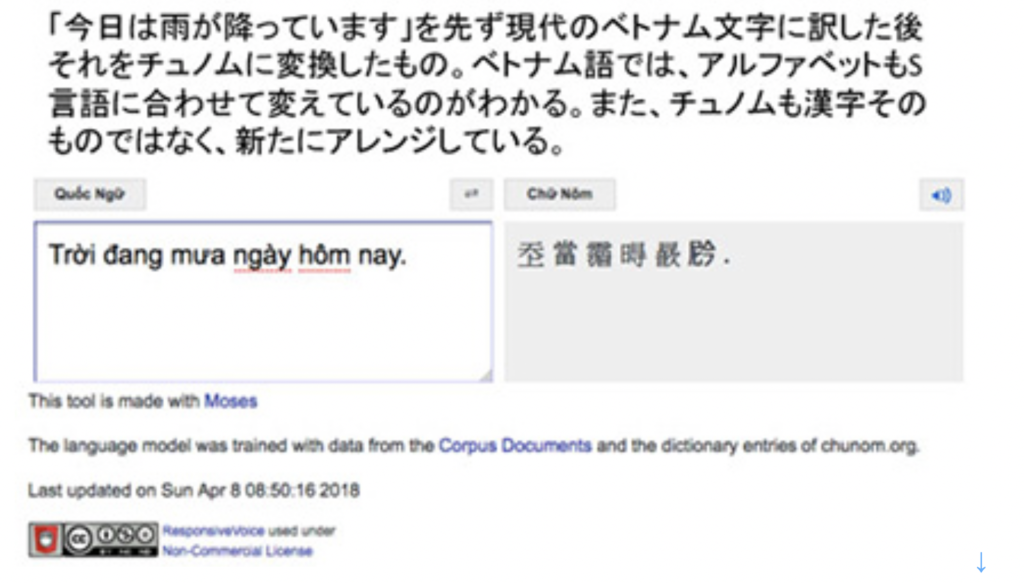
図1 現代ベトナム語と、チュノム:Google翻訳で日本語の「今日は雨が降っています」を現代ベトナム語に変換した後、上記のアプリでベトナム語をチュノムに変換した。
S言語も社会と個人を行ったり来たりする間に進化を遂げるが、文字も使う人が増えれば増えるほど拡張し変化していく。日本語を見ながらこの点を整理してみよう。
日本語では今も、中国で誕生した表意文字(logogram)漢字をほぼ原型に近い形で使っており、常用漢字が約2000字存在する。しかしこれ以外にも普通に使われる漢字は存在し、一般の人でも辞書なしで読める漢字の数はかなりあるのではと思う。さらに、ワープロが使われるようになってからは、個人の記憶の制限がなくなり、50000字近い漢字が比較的容易に使えるようになってきている。おそらく、日本語の場合、ワープロのおかげで今後も漢字の利用は拡大するのではと予想する。
日本語での漢字の使い方で重要なのは、中国語起源の読みと、日本語読みを漢字に当てた訓読みが両立していることだ。このことから表意文字は、意味が合えば文字をどう読んでもいいことがわかる。このおかげで、中国語と日本語が合体した新しい単語を作ることすらできる。日本語の場合、熟語の多くは音読みが普通だが、例えば「台所」のように「台:ダイ」という音読みと、「所:ドコロ」という訓読みを合わせて一つの単語が作られる。ちなみに中国語で台所は「厨房」(チューファン:チュウボウ)なので、台所は日本の造語だ。
音読みと訓読みを両立させることで同じ単語の持つ違うイメージを表現することもできる。例えば「身体」を「シンタイ」とよむとよりアカデミックに聞こえるし、「からだ」とよむとより身近に感じる。面白いのは、訓読みの「こころ」に対応するときは「からだ」と読むし、「精神」と音読みに対応させるときは「シンタイ」になる。日本語を初めて習う人から見ると大変難しい部分だが、一つの単語を漢字で表すことで、様々な多義性を表現できることは、表現力という点では優れている。現在中国で漢字がどのように使われているのか悲しいかな把握できていないが、私は漢字の持つ能力を最大限に生かしているのは日本語ではないかと思っている。
このように、漢字のポテンシャルを最大限に利用できるのも、日本語が漢字をもとに、ひらかな、カタカナという2種類の表音文字を開発し、現在も両者を区別して使っているからだ。表意文字から表音文字ができる歴史は後回しにして、現在使われているカナについて見てみよう。今、「仮名」を表音文字と言ったが、2種類の表音文字を両立させることで、表音文字に一定の表意性を持たせることに成功している。例えば、外来語はカタカナで書くことが多いため、カタカナの単語は外来語や音の表記であるというカテゴリーを示すことができる。
一方、この節の最初のセンテンス
「このように、漢字のポテンシャルを最大限に利用できるのも、日本語が漢字を元にひらかな、カタカナ….」
を見てわかるように、私たちは平仮名を一つの単語を表すのにほとんど使わず、単語をつなぐ助詞や、接続詞に使っている。一方、カタカナは現在ではもっぱら単語に使うようになっている。2種類も表音文字を持つのは大変だが、表音文字の最大の欠点である、単語のカテゴリーの表現ができないという問題が、日本語では表音文字でも解決されている。
現代の日本語を見ると、さらに多くの文字が使われている。数学的な記号はほぼ完全に西欧の表記を使っている。数字に関して、もはや縦書きの文章でもアラビア数字が使われることの方が多くなり、漢数字使用頻度は低下の一方だ。他にも、+、-、%などはほぼ100%西欧から輸入した文字を取り入れている。ちなみに、これらは全て表意文字に分類される(ということはアルファベットを使う国でも表意文字を使っていることになる)。さらに、私が今書いている文章もそうだが、必要に応じてアルファベットも平気で使うし、メールに至っては新しい表意文字といえる絵文字も使われている(ちなみにEmojiは世界標準の単語になっている)。おそらく、外から見るとどうしてこんな複雑な文字体系が滅びなかったのか、不思議に思えるだろう。
漢字と2種類の仮名を使うということが、日本語の文字の節操がないとすら思える拡張性を支えている。漢字の多くを音読みすると述べたが、これは単語の多くが中国から入ってきたからだ。仮名が開発された後は、こうした輸入単語をそのままオリジナルな文字で書かれた単語として文章の中に置くだけでいい。たとえば、「これからBaseballを見に行こう」といった感じだ。ただ、中国以外からも様々な単語が入ってくる時代が始まる。単語を漢字で表現する伝統を守っていた日本語でも、ヨーロッパからの言葉も、最初は一度漢字に当てはめて使用されていた。
面白いことに、本家の中国語では、ヨーロッパ人の宣教師たち自らが、ヨーロッパの言葉を、漢字に訳し直したことが知られている。わが国でもこの時漢語に訳し直されたヨーロッパ由来の単語を中国から輸入しそのまま使っている。たとえば、「基督」「天主」などキリスト教の単語はもちろん、「地球」「医学」など、現在も広く使われる単語がそれにあたる。この漢語に書き直す伝統は、わが国に直接入ってきたヨーロッパ語にも最初適用される。ただ、日本では外来語をそのままの音を使うことに抵抗は少なく、padreを伴天連、Christianを切支丹と表記することになる。
もちろん我が国でも、意味を訳し直すことも行われた。最初に行われた最も有名な翻訳作業の例がオランダの解剖学の教科書「ターフェルアナトミア」の翻訳で、それぞれをどのように訳し直すかが最も苦労したことが杉田玄白の蘭学事始めに書くかれている。ただ、こうしてできた和製単語は中国でも利用されるようになり、ここでも表意文字の普遍性がわかる(図2)。
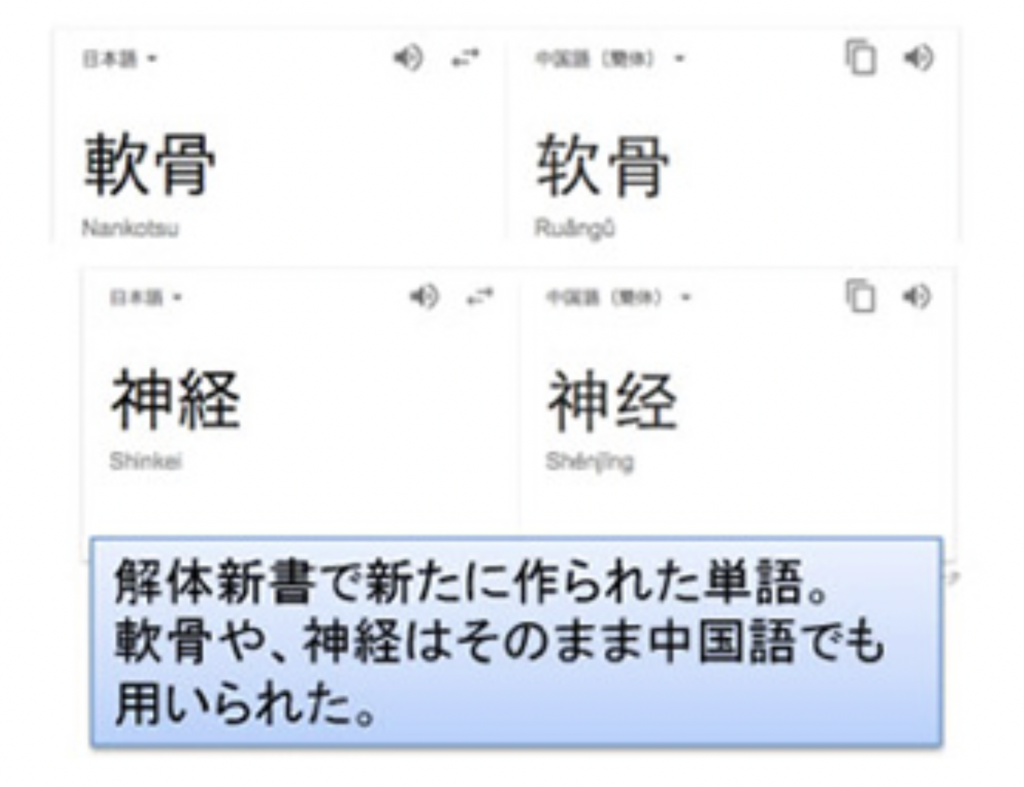
図2 解体新書で漢語に訳し直された単語と、現在の中国語での表記。Google 翻訳で訳した。中国語では漢字が簡略化されている。
ただ、このように単語の音ではなく、意味を取り出し自国語に訳し直すのは大変な努力が必要になる。幸い、日本語ではカタカナという伝家の宝刀が利用できる。その結果、難しい単語は中国起源であれ、日本起源であれカタカナ表記に変えることができる。今は中国由来の言葉「基督教」は「キリスト教」にほぼ完全になっているし、わが国由来の訳語「煙草」(ちなみに中国語では烟草)は「タバコ」に変わっている。このおかげで、私たちはIce creamやcakeをアイスクリーム,ケーキと、かって漢語をそのまま輸入したように簡単に日本語の中に導入できる(ちなみに中国語ではそれぞれ「冰淇淋」「蛋糕」となる)。
現在の日本語の特徴を見てきたが、この節操のないと言っていい拡張性には幾つかの秘密があるようだ。
一つは、表音文字を2種類持つことで、表音文字に表意性(カテゴリーについて)を加え、外来語であることを明示しながら、そのまま外来語の音を表記するのに成功した点だ。もともと日本語は漢語として様々な単語をそのまま導入することに慣れていた言語で、外来語をカタカナというカテゴリーを示すことのできる表音文字に任せることで、文章がわかりやすく整理できる。たとえば、「わたしはあいすくりーむを、すぷーんでたべます(我用勺子吃冰淇淋)」と書いてもわからないではないが、「私はアイスクリームをスプーンで食べます」のほうが意味をとるのにかかる時間は、中国語と比べても短くて済むと思う。おそらく、この構造がわが国では文字を読むことに異常をもつ「失読症」が少ない理由ではないかと想像される。
もう一つの秘密は、日本語が、単語自体が変化して格や時制を表す屈折語ではなく(例えばドイツ語の愛する、liebe, lieben, liebst, liebtなど)、助詞を単語の間につなぐ膠着語であるという点だ。この屈折語は、中国語のような孤立語、すなわち単語を並べるだけの言語の文字に極めて相性がいい。すなわち、単語をつなぐ役割の助詞などにひらかなを用いるだけで、何の苦労もなく外来の単語を利用できる。ただ、昔と違って私たちはこれを平仮名に任すことに慣れている。そのため、現在では助詞などがカタカナで表記されると、途端に文章の理解が送れるように思う。逆に電報文「チチキトク(父危篤)」のように、カタカナだけの文章で助詞を使わないで短く表現していることを明確に示すことができる。
膠着語としての日本語を理解した上で、日本語が漢字を導入する前から膠着語だったのかという問題はとても面白い問題だ。残念ながら、古代に話されていたS日本語について教えてくれる資料は全くない。そのためここからは私の個人的な妄想だが、日本語は漢語のような格や時制すらない言語から生まれた表意文字を導入したことで、外来の単語を日本語でつなぐという膠着語を完成させたように思える。これまで議論してきた日本語の特徴が全てを、高校で習った漢文に見ることができるが、この杜甫の詩を見ていると漢字を使うことで日本語が膠着語を完成させたような気にますますなってくる。

図3 杜甫の漢詩。音読み訓読みが混じり、それらが助詞などで繋がれている日本語の特徴がよくわかる。
お隣の韓国も、現在は漢字を使わないが、昔は日本語と同じで文字は漢字に頼っていた。そして、やはり文法的には膠着語だ。ちなみに、ベトナム語は孤立語で、チュノムの使われ方を知ると、これも納得できる。
もともと、表意文字は孤立語的傾向を持っている。これをそのまま使い発展させた中国語は例外で、次回以降紹介するメソポタミアやエジプトでも、絵文字のような表意文字は、その後それから表音文字を開発し、両方を使うようになるが、この時、言語は膠着語のような特徴を持ったようだ。その意味で、私は日本語の膠着語性は、漢語を文字として使うことで完成して行ったのではと信じている。
既に述べたが、漢字が導入される前に日本語が文字を持っていたかどうかはわからない。ただ、文字の原型と言える視覚を用いるコミュニケーション手段はおそらく直立原人の頃から存在していたはずで、その意味で日本語に独立した文字が存在しなかったはずはないと思っている。ただ、日本語の場合、オリジナルな文字は、S言語と一体化して、新しい言語を作るにはいたらなかった。そのかわり、4世紀朝鮮遠征を機に多くの朝鮮文化人がわが国に渡来したこと、そして仏教の渡来ととともに多くの経典がわが国にもたらされたことで、外来語ではあっても漢字が文字として選ばれる。幸いにも、漢語が表意文字を用いる孤立語であったことで、文字としては普遍性が高く、しかも音読みと訓読みを並立させることで抵抗なく文字としての漢語を受け入れることができたと思われる。
とは言っても、漢字がS言語と一体化するためには、日本語の本来持っている文法構造に合わせ、また漢語にはない多くの単語を表現する必要がある。これは表意文字の宿命とも言える過程だが、よく使う表意文字を、その読みの発音だけを表現する文字への転換が起こる。
日本語の場合、これが万葉仮名になる。万葉集で使われたことから(図4)万葉仮名と呼ばれているが、実際には漢字が導入されて間もなく、音を名前などに当てはめて使われている。このことから、表意文字が一定のレベルに達すると、文字は表音文字としてすぐに利用されるようになることがわかる。

図4 万葉仮名で書かれた額田王の有名な和歌。
使われている仮名は、迩(に)、世武登(せむと)、者(ば)、毛(も)、可奈比沼(かなひぬ)、者許芸乞菜(こぎでな)(写真出典:Wikipedia)
万葉仮名の場合、一つの音に対して幾つかの漢字が当てられる。そしてこの表記の複雑性を単純化して統一するために仮名が登場するが、仮名が完全に整備されるのは平安時代以降のことになる。
図5、ひらかなとカタカナの起源
出典:Wikipedia
なぜカタカナとひらかなが同時に生まれたのか私には想像もつかない。それぞれの表音文字は、一部を除いて別々の漢字に由来している。ただそれぞれが使われている資料として残っているのは、ひらかなが歌などの文学、カタカナは漢文を読むときの注釈として使われ、公的な文書に使われたのが記録として残っている。この違いが、現代での両者の使われ方の違いにつながっていると思うが、これ以上探求はしない。しかし、2種類の表音文字を持ったことが、私は日本語の拡張性に大きな影響を持つことになったと確信している。
以上日本語の文字の成立から、
ことがわかる。次回からは、メソポタミア、エジプトの文字を例に、文字の変遷を見ることにする。
[ 西川 伸一 ]
2018年5月1日
前回は絵文字から表意文字、表意文字からRebusとよばれる表意文字の表音化(万葉仮名やベトナムのチュノムにあたる)、そして表音文字の誕生と、紀元前3000年頃以降エジプトやメソポタミアで起こった文字の発達過程の全てが、日本語では今も生きて使われていることを説明した。今回から視野を世界規模に広げ、文字誕生の過程を説明する。
音節やジェスチャーに依存した言語の始まりと違って、文字は遺物として残されるチャンスが高くエビデンスに裏付けられた話をすることができる。これまで出土した遺物で見る限り、絵文字も含めて、文字と呼べるものは早くても紀元前3000−4000年頃に始まったと考えられる。音節によるS言語誕生時期について特定されているわけではないが5万年前後とすると、文字が誕生するまでかなり時間がかかったことになる。S言語誕生から文字誕生まで、人類は農業や離れた集団間の交易を始め、この過程で個人の貧富の格差がはっきりと見える国家が形成されていった。事実、エジプトのヒエログリフや、メソポタミアの楔形文字、そして中国の漢字など、出土する場所から考えても、文字が国家の形成とともに発生したことは間違いない。
文字は様々な機能を持つが、最も重要なのは長期間維持できる記録だと考えると、国家のような複雑な人間関係に基づく社会の形勢に文字が必要なのは直感的にもよくわかる。従って、文字の歴史を考えるうえで最も重要なことは、人類進化のどのような状況が記録を必要としたかを考えることだ。
さて、前回日本語で使われる文字のルーツ漢字が絵文字から生まれた漢字であることを強調したため、全ての文字の起源は絵文字であるような印象を与えたかもしれない。実際、漢字だけでなく、エジプトのヒエログリフやインカの文字をみると、これらが絵文字から始まっていることは間違いない。このため、メソポタミア地方での発掘が進むまで、文字の紀元は絵文字だと考える人は多かった。
S言語とともに、人類は絵をコミュニケーションの手段に使う能力を持っていたことは、ラスコー(2万年前)やアルタミラ(1万8千年)の洞窟に残された絵の素晴らしさを見ればわかる(図1)。このような絵を、より使いやすい形に変形して絵文字が生まれ最初の文字になったという考え方だ。
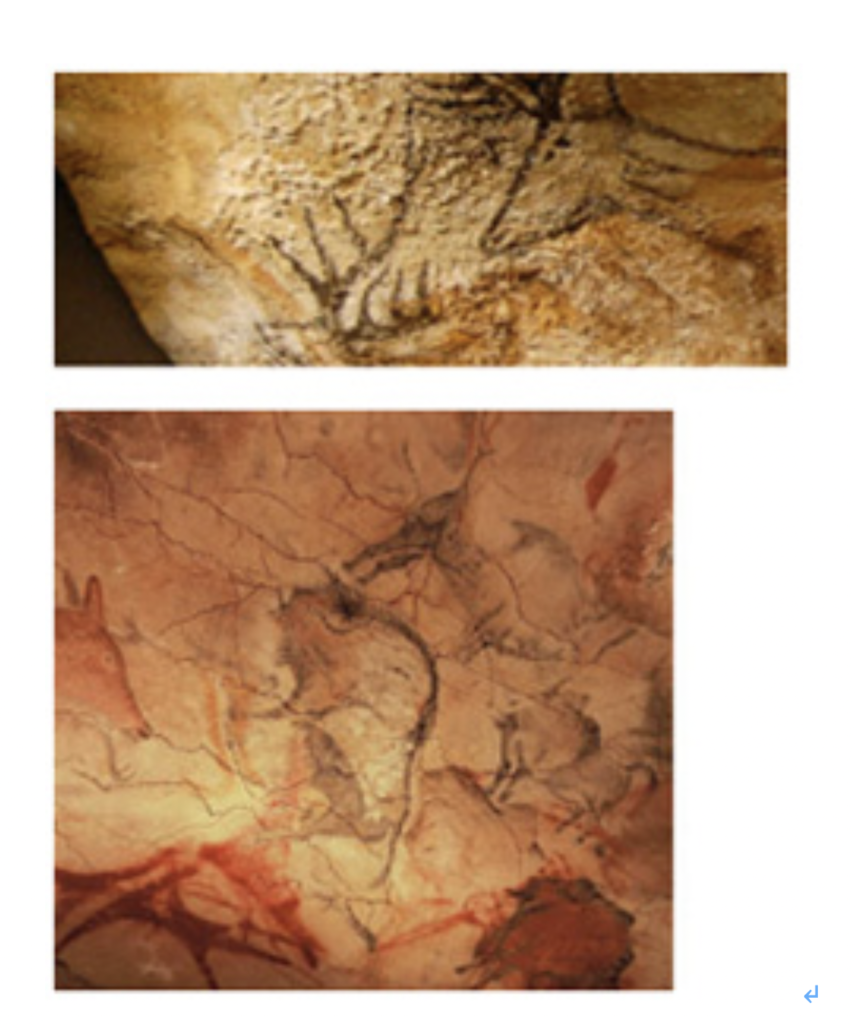
図1:2万年ごろの現生人類により描かれた鹿の絵。上がフランスラスコー洞窟、下がスペインアルタミラ洞窟。(写真出典:Wikipedia)
しかし現在この考えを持つ言語学者は少ない。というのも、最初に文字が発生したメソポタミアで、エジプトの代表的絵文字ヒエログリフより古い時代の遺物をしらべても、エジプトのように絵文字だけで書かれたタブレットを発見することが難しいからだ。もちろん絵文字も存在するが、文章の一部に使われる程度であることがわかっている。このことから、記録が最初に必要とされる状況では、絵を用いて表現するよりもっと簡便なアイテムの表現方法が必要とされていたことを示唆している。この絵よりも先に利用されていた記録が、今日紹介したいと思っているトークン(Token)だ。
さてトークンとは何か?私自身トークンと聞いて何が頭に浮かぶかと問われれば、ニューヨークの地下鉄で切符代わりに使われていた代用通貨のことを思い出す(図2)。現在ではすでにメトロカードに置き換わっているので、おそらく若い方は聞いたこともないかもしれない。

図2 ニューヨーク地下鉄で切符代わりにつかわれるトークン
私の世代が、トークンと聞くと最初に思い浮かべる、NY メトロで使われていたトークン。何かの代用をするという意味で使われるもので、今日紹介するトークンも何かのアイテムを代用しているが、アイテム特異的で、あらゆるアイテムを代表できる通貨とは違っている。(写真出典:Wikipedia)
今日紹介するトークンとは、BC8千年以上前(まだ文字が使われているという証拠のない時代)から、文字が日常使われるようになる時代まで、最初メソポタミアで、その後様々な地域で使われていた、農産物など、生産されたアイテムを数え記録する時に使われた一種の道具のことだ。トークンに関する私の知識はトークン研究の第一人者デニス シュマント=ベッセラ(Denise Schmandt‐Besserat)の著書『How writing came about』(University Texas Press)から仕入れたもので(図3)、訳本はないがこの分野に興味のある方はぜひ読んでほしい本だ。

図3 今回の原稿を書くにあたった最も参考にしたSchmandt-Besseratの『How Writing Came
About』。(写真出典:Amazon.co.jp)
巻末に様々な時代に、様々な場所から出土したトークンを網羅した図鑑が掲載されており、トークンのことを知るには最適の一冊。
この表紙の写真に示されているのが、最も古いトークンと、それをしまうエンベロップと呼ばれるコンテナーで、たとえば丸いトークンが酒一升、四角いトークンが米一俵といった具合に考えて貰えばいい。
ここでちょっとみなさんがものをどう数えているか思い出してほしい。たとえば、今部屋に何脚椅子があると聞かれたとする。今これを書いている顧問室には、総計8脚の椅子があり、数えなくとも頭に入っている。しかし初めての部屋だと「一、二、三….」と指差しながら心でつぶやいて数えるはずだ。ただ、これは私たちが言葉としての数字を使い慣れていないとできないことは明らかだ。もし全く数字の概念がなかったら、どう勘定すればいいのだろう?
「そんなことはない、文字を持つ前から数の概念は必ず存在したはずだ」と言われるかもしれないが、数の概念は抽象的で、よほど必要にかられないと、数を正確に数えることが必要ないことは、原始的生活をおくっている少数民族の言語を見るとよく理解できる。最も有名なのはスリランカの少数民族Wedda(Vedda:図4)で、彼らは、one, twoの次にはmanyしか数の概念がなく、4本でも5本でも、たとえば木の数は、manyと表現されてしまう。

図4 スリランカのWedda
2以上の数の概念がないことで有名な少数民族。
(写真出典:Wikipedia)
おそらく我々文明人でも同じで、数字に関する言語的表現を完全にマスターしていないと数を数えるのは難しいことは、幼児を観察しておればわかる。ましてや、せっかく苦労して手に入れた穀物の量を数えるだけでなく、それを長期間記録することは、文字を持たない時代にはほとんど不可能だったと推察できる。
チグリス川、ユーフラテス川に囲まれた肥沃な三日月地域とその周辺では、少なくとも1万年以上前、文字のない時代から農耕が行われていたことは間違いがない。農業は、長期間保存が必要な穀物を生み出す。また、農業を支えるためには、様々な道具やインフラ、さらには狩猟採取民の生活よりはるかに複雑な人間関係が必要で、これらの活動を支えるために、数え、記録することが必要になったことは想像に難くない。
ではこの課題を、文字や数字のない時代にどう解決したのか?その切り札として登場したのがトークンというわけだ。粘土で簡単な形を作り干して作られた小さな陶器で、特定の形のトークンを、数を数えて記録したいアイテムに対応させる。そして、アイテムの数に合わせたトークンの数をまとめてどこかにしまっておくことで、そのアイテムを今いくつ持っているのか記録するという仕組みだ。たとえば通貨のように交易に使われたものではなく、あくまでも個人や集団の記録として利用されたと考えられている。
同じような目的に使われる他の記録方法には、ノッチと呼ばれる動物の骨や木切に切れ目を入れて数字を記録する方法や、Tallyとよばれる線を足していく方法がある。
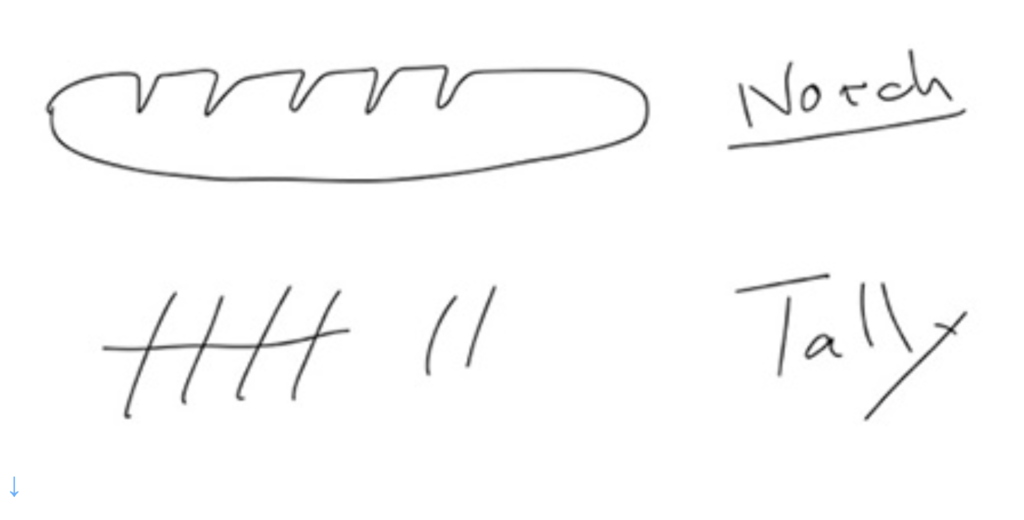
図5 NotchとTally
しかしこの方法だと、記録するためのアイテムの数が増えると簡単には対応ができない。実際社会が複雑になり、様々なものが生産されるようになると、単多くのアイテムを数える必要が出てくる。その点トークンだとその形を複雑にしたり、あるいはトークンの中に図を入れることが可能だ(図6)。

図6:メソポタミアUruk時代後期(BC3300年ごろ)に使われていた様々なトークン。すでに、この時代には文字も使われ始めているが、トークンも併用して使われていることがわかる。(NYメトロポリタン美術館で筆者撮影)。
トークンが使い始められるのはBC8000年頃と推定されているが、最初は数えるアイテムが少なかったためか、図3や図6の両端にあるような単純な形のトークンだけが使われていた。トークンが開発されて以降、ほぼ5千年近くはこのような単純な形のトークンのみが使い続けられていたことから、農耕社会ではそれほど多くのアイテムの記録の必要がなかったことがわかる。ところが、BC3000年頃のUruk時代のように、都市が形成されだすとそこで作られるアイテムの数は増大していく。その結果、図6で示すような様々な形のトークンが開発されていく。
以上、トークンとは何かについてかなりお判りいただいたと思うが、まとめておくと以下のようになるだろう。
トークンは当時の人たちにはとても便利な方法だったようで、メソポタミアだけでなくエジプトをはじめとする他の文明にも伝えられていく。そして、伝えられた先々で、文字の誕生を促したのではと考えられるが、それについては次回に回す。
[ 西川 伸一 ]
2018年5月15日
最も古い文字は絵文字(picotgram)だっただろうと推定できるが、文字として使われていたことが間違いないと判定できる絵文字が見つかるのはBC3−4000年前からで、それ以前には絵は存在しても、絵文字は存在しなかったと考えられている。発見が難しいからこれより古い文字が見つっていないだけだという意見もあるが、本当に存在しなかったと言える証拠の一つが前回紹介したトークンだ。トークンはBC8000年前の遺跡にはその存在が確認されているが、その後BC3-4000年に文字が使われ始めた時代にもまだトークンは文字と一緒に使われていた。すなわち、トークンが使われていた約5000年間、もし文字も使われていたとすれば、必ずトークンと共に遺物として発見されるはずだ。それが全く発見されないということは、トークンが使われていても、文字は使われていなかった時代が5000年近く続いたことを示している。
ではトークンとはなんだったのか、もう一度おさらいしてみよう。まず、トークンは農耕が普及し、穀物を貯蔵しその量を記録する個人的必要から始まったと考えられている。すなわち、一つのアイテム(例えば米一俵、酒一升など)を一つのトークンに対応させ、現在何俵の米を所有しているのかを記録するために用いられた。貯蔵した穀物を使い果たすと、そのトークンは廃棄され、次の収穫には新しいトークンが使われた。この点で、繰り返し交換に使われる通貨とは全く違っている。前回述べたが、文字や数の抽象的概念を使いこなすようになる以前は、数を数えて記録することは簡単ではない。その意味で、トークンは大きな発明だったと思う。
トークンの数を表現するのに、抽象的な数の概念を表すサインが使われることはなく、対応するアイテムの数だけトークンが利用されたことも重要だ。米3俵に対応するトークン3個を記録として残す場合、一個のトークンは米一俵という具体的なアイテムに対応するため、記録には3個のトークンが必要になる。絵文字でみかん2個を🍊🍊のように表現するのと同じだ。
こう考えると、トークンは具体的なアイテム(例えば米一俵)をシンボル化しているという点で、すでに言語における単語の役割を果たしていることがわかる。もちろん、各タイプのトークンには話し言葉の音節も対応していたはずで、この点ですでにS 言語に対応する文字として考えることも可能だ。ただ、開発されて以降、長期にわたって、トークンは独立して個人的に利用されるだけで、文章の一部として使われることはなかった。いわば、赤ちゃんが最初「ママ・ママ」と単語だけを叫ぶ段階と同じだ。しかし話し言葉で表現される単語に対応するサインとして使われていることは間違いなく、話し言葉を文字化する先駆けになっている。
すでに言語を話している人間が、トークンとして現れた単語に対応する文字の原型を文章の中に組み込み始めるためには、「XXが」「トークン」を「XXした」というセンテンスを文字として残す必要性が発生するまで待たなければならなかった。個人のメモがわりに使っている段階では、このような必要性はなく、所有関係や交易が複雑化した社会構造が生まれたとき、トークンが文章の一部に埋め込まれだす。この意味で、文字と文章、すなわちWritingの始まりに、トークンが触媒として大きな役割を果たしたと考えられている。
前回、トークンはコンテナーに入れたり(前回の図3に掲載したBesseratの表紙にトークンと共に示されている入れ物)、後期になると紐に通してしまって置かれたことを紹介した。紐を通してしまう場合は、トークンの数がはっきりと見えるので問題はないが、コンテナーにしまう場合、図からもわかるように外からいくつ入れたのか見ることができない。そこで、粘土が柔らかいうちにトークンで刺したり、押し付けたりして、トークンの数が外から見られるようにする方法が開発される(図1)。すなわち、トークンが一つの平面上に表現され始めたことを意味し、これにより一歩文字に近づいたと言える。

図1:トークンとそれをしまっておいたBullaと呼ばれるコンテナー。
Bullaの表面に、トークンで穴が開けられて、幾つのトークンが入っているのか確認することができる。(写真出典:FinalySchool)
しかしこの段階でもまだ文章とは言えない。もちろん個人のメモ書きであればこれで十分だ。しかし、社会が複雑になってくると、収穫が全て自分の所有物とは言えなくなる。例えば宗教が発達すると、収穫は神社に供出され、管理される。さらには、社会が階層的になると、支配者に一部の収穫が収められ、残りを自分で管理するといった状況が生まれる。すなわち、誰が収穫を所有するかを明記することが重要になる。この時、所有している主語のサインがトークンと共に使われるようになると、文章のための文字が誕生したことになる。
図2は、メソポタミアの楔形文字が生まれる以前の文字が書かれたタブレットだが、上部にはトークンを押し付けた跡が示されているが、収穫物の分配が記録されていると考えられる。

図2:ルーブル美術館所蔵の楔形文字の原型になった絵文字の書かれたタブレット。絵文字には、召使いの名前が書かれていると考えられている。上の穴はトークンを押し付けて作ったと考えられる。(出典:Wikimedia Commons)
この結果、誰がトークンに表現されているアイテムを持っているのか、あるいは生産したのか、などが文章として表現されていく。すなわち、個人的なメモとしてのトークンから、所有や生産を主張するためのトークンへと変化する過程で、所有者の名前を特定する絵文字が加えられるようになった。繰り返すと、社会の変化が、文章を文字で表すことを促したが、この時この触媒として働いたのが、所有の記録としてのトークンということになる。前回紹介したBesseratの本には数多くのトークンの形が紹介されている。社会が複雑になるに従い、記録に必要なアイテムは増え、それに対して形態や模様の異なるトークンが作られていったことがわかる。さらに彼の本には、トークンが粘土板に押し付けられて描かれたサインの形のリストも示されている。実際には、18種類のトークンと、それを押し付けられて作られたサインが示されているので、これだけでも18種類の単語が存在したことと同じになる。
しかし、押し付けるだけでタブレットに記録できるトークンは単純な形態に限られる。ところが、都市が発達してトークンの使用が農業以外に拡大するに連れ、トークンの形態は爆発的に多様化し、複雑になる。実際、ミルク、ビール、犬、羊、ロープなどとありとあらゆるものに対応してトークンが作られる。このようなトークンは粘土に押し付けるだけでは正確にコピーすることができない。結局、それぞれのアイテムを押し付けるのではなく、おそらくBullaやタブレットに直接トークンの形をペンで書き入れるようになったと考えられる。実際、Bessartの本では、トークンに由来する多くの絵文字が紹介されており、生き生きとしたトークン文化を知ることができる。図3は、彼の本に掲載された図の一部を拙いスケッチでコピーした図だが、ビール、ミルク、ロープなどに対応していたトークンが、そのまま絵文字へ転換されたことがよくわかる。
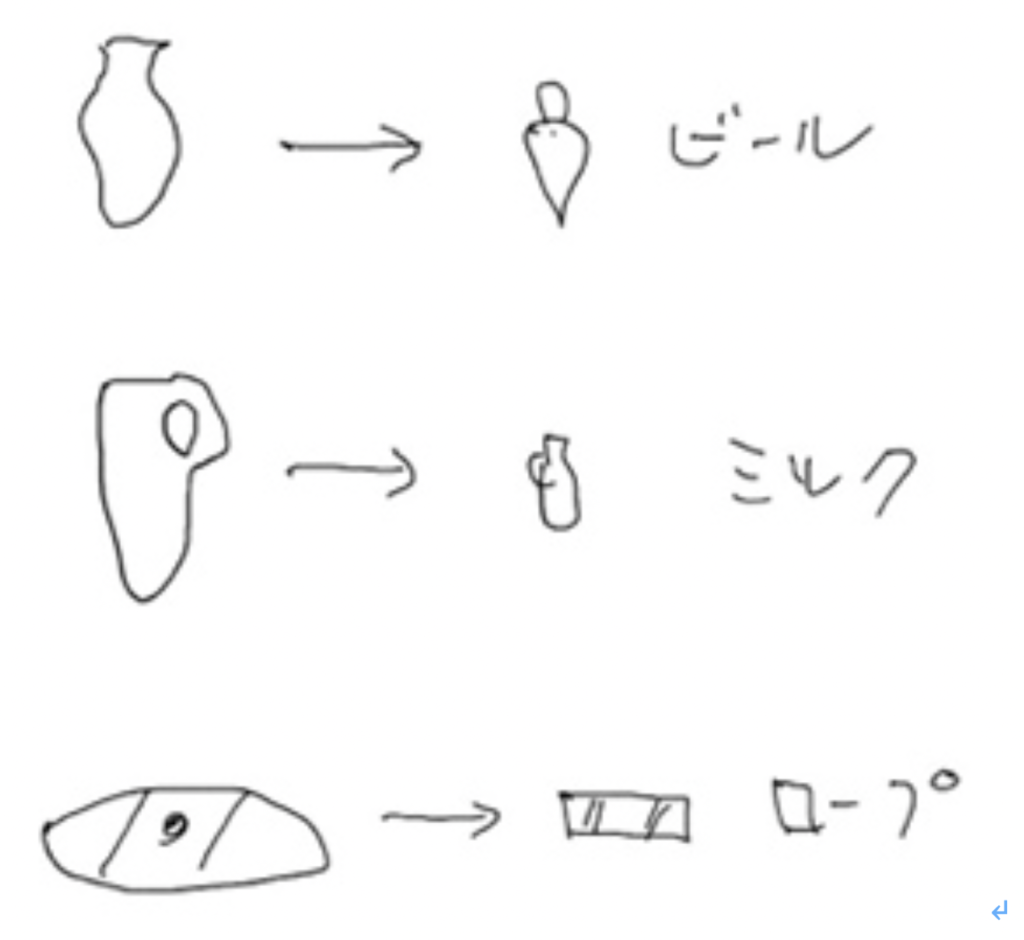
図3 左がトークンで、右がそれに対応する絵文字
Besseratの著書より抜き書き(例としてあげただけで極めて不正確)
ここまでくると、Writingの完成も近く、あとはトークンとは独立に、絵文字のボキャブラリーを増やし、あとは主語や述語を絵文字として足していけばいい。もともと、文章は話し言葉として使い慣れている。それを文字として残すことができることが理解されれば、Writingは誕生する。これは私の勝手な想像だが、図2にある手のサインは、「持つ、所有する」という動詞に変化したのではないだろうか。
以上、絵文字や文章が、トークンの使用をきっかけに、5000年近くかかって生まれたことがおわかりいただけたと思う。次回は、絵文字から表音文字への変化を見ることにする。
[ 西川 伸一 ]
2018年6月1日
今回以降、「文字」と表現する代わりに「Writing」という単語を使い、「書き現わす」技術と言うと広い意味の中で文字の誕生について考えていく。
Writingは最初絵文字(Pictogram)から始まり、その起源は絵を描く人間独自の能力に起因するという考えがある。直感的には説得力があるが、まず2回にわたってトークンの話を続けたのは、Writingが絵から絵文字へ進化する延長線上に誕生したのではないことを理解してもらうためだ。すでに見たように、トークンはまず自分自身の覚え書き的メモとして使われ始めた道具が、タブレット上の記録へと変化し、最後に絵文字や表音文字も加わって文章をサインで表すWritingへと発展する。しかしそんな説明をしている私自身も文字について学ぶまではWritingは絵を描くことから始まると思っていた。この考えを改めてくれたのがバリー・B・パウエル(Barry B Powell)の『Writing: Theory and History of the Technology of Civilization』(Wiley-Blackwell)で図1に示す。

図1 Barry B Powellが2009年に出版したWriting についての著書。
Barry Powellは1942年生まれのギリシャ文学史研究者で、ギリシャについては「オデッセィ」など多くの本を出版している。
ここに紹介する彼の著書は、Writingの歴史や理論について知るためには極めて優れた著書で、ギリシャだけでなく、文学に対する深い教養に裏打ちされており、大変わかりやすい。
トークンが確かにWritingの起源の一つであることはわかるが、ではトークンの延長に現れる絵文字がなぜ絵画と関係がないと断定できるのだろうか?事実、精緻な絵は、話し言葉が使われだしたと考えられる3−5万年前から描かれてきたことは多くの証拠により裏付けられている。洞窟に壁画として残された絵は、石器時代に描かれたとはいえ、今日上手な画家が描いた絵画といっても何の不思議もないレベルだ。おそらく多くの人は、スペインラスコー洞窟の壁画や、世界最古の洞窟壁画と言われているフランス・ショーベ洞窟に残されたこのような精緻な壁画の写真を一度は見たことがあるのではないだろうか(図2)。

図2 ショーベ洞窟の壁画を再現した絵 出典:Wikipedia
図2に示した絵が描かれたのは今から30000年以上前だと知ると、人間は早くからリアリズムに満ちた絵を描く能力を持っていたことがよくわかる。とは言え、このような素晴らしい絵を当時のほとんどの人間に描く能力があったのか問うてみると、答えはまず間違いなくNOだと思う。事実、現代の私たちでも、このような真に迫った絵をかける人は限られている。まず私には不可能だ。即ち、ここまでの絵を描ける力は当時も今も、一部の人に限られている。
ちょっと脱線するが、自閉症を考古学の立場から考えている若手研究者ペニー・スパイキンズ(Penny Spikins)は最近発表した論文(How do we explain autistic traits in European upper palaeolithic art, Open Archeology: https://www.degruyter.com/view/j/opar.2018.4.issue-1/opar-2018-0016/opar-2018-0016.xml)やオンラインブック(The prehistory of autism: https://roundedglobe.com/books/850efa8a-9596-44c5-ab46-5dd828905986/The%20Prehistory%20of%20Autism/)で、これら旧石器時代の目を見張る絵画やフィギャーは、現在自閉症スペクトラムの人たちがしばしば示す、local processing bias(全体ではなく特定の領域にこだわって視覚的に認識しそれを表現できる能力)を備えた限られた人たちにより描かれたと推察している。彼女は、現在ではともすると精神発達の異常として排除される自閉症スペクトラムが、石器時代では社会にとって得がたい能力を持つ人間として重宝されていたこと、そしてその結果、自閉症スペクトラムと連鎖している何百もの遺伝子が、人類の遺伝子財産として一部の人に現在まで受け継がれてきたと考えている。図3は2015年に出版された『How compassion made us human(思いやりはどのように私たちを人間にしたのか?)』だが、彼女の研究方向がよく分かる良書だ。
図3:Penny Spikinsがおそらく30代で出版した著書。優しさや、道徳性など人類の良い部分の起源について考古学的に考察した本。(出典:Amazon.co.jp)
私はこの本を読んで以来、彼女の大胆な説に賛同している一人だ。彼女の考えと、自閉症スペクトラムの人の症状、すなわち社会との付き合いが下手なのと同時に、言葉の発達が遅れるという症状考え合わせると、たしかに優れた絵画を描く能力は、言語とは全く独立していたように思える。彼女が示唆するように、このような空間認識力に長けた写実的絵画を描くために、local processing biasをもった絵画能力が必要なら、絵を描くという行為が決して一般的なコミュニケーション手段にはなれないことがわかる。言語はすべての人が話す媒体でなければ役に立たない。このことから、Writingとショーベの洞窟に残るような絵画は、共通性はあっても、因果的な関係はないと考えられる。
繰り返すが、言語がコミュニケーションの手段である限り、その使用が限られた人に限定されると言語になり得ない。しかし前回Writingの先駆けではないかと紹介したトークンのような覚書のための道具は、道具であるがゆえに社会全体で共有することができる。と言っても、トークン自体は道具であり、それ自体は話し言葉とは全く独立し、文章の中に使われるには何千年もの時間が必要だった。同様に、誰でも描くことができる原始的な絵によるサインも、話し言葉とは独立して使われていたように思える。例えば絵文字で(☀→☔)と書いたとき、「晴れ後雨」でも「clear then rain」でも読み方とは独立して意味を示すことができる。おそらく早くから、このようなサインは使われていたのではないだろうか。例えば何かを教える時に、地面に枝で簡単な絵を描いていたのではないだろうか。このようなサインはメモ書きのようなものなので、洞窟絵画のように丁寧に描かれることはなかった結果だと想像する他、本当にこのようなサインが使われたのかどうかを知る証拠は残念ながらほとんど保存されていない。
トークンも、原始的な絵を使ったサインも話し言葉とは独立していたとすると、Writingの誕生には、話し言葉の文章を、独立した書かれたサインと、音節を媒体にするS言語を融合させることがまず必要だった。おそらくWritingの誕生は、自分の話しているS言語の単語に対応するサインを順番に作成することで起こったのではなく(必要性が認識されてもサインを決めていくのは大変な作業だ)、独立して存在していたS言語とトークンなどのサインがたまたま融合する過程で新たな使い方が着想され、さらに新しいサインを生み出していったと考えられる。
トークンでも、トークン由来の様々な絵文字(前回図3)でも、あるいは誰でも簡単に描ける絵を基礎にした簡単なサインでも、もちろんそれに対応する発音は存在し、そのサインを見てその単語が発音されていたのは間違いない。このように、サイン自体に意味が存在し、話し言葉とは独立している文字はLogogramと名付けられている。日本語では漢字がそれにあたる。ほとんどの漢字はそれ自体で意味を表現しており、音読み(中国語)でも訓読み(日本語)でも可能で、漢字を理解する人なら英語で読むことすらできる。同じことは世界中で使われているlogogram「1、2、3」を「ichi, ni, san」「Yi, er, san 」「one, two, three」「ein, zwei, drei」と違った読み方ができるのと同じだ。すなわち漢字は話し言葉の発音とは独立している点でlogogramと言える。しかし日本語の文章をlogogramだけで表現しようとすると難しい。どこかで、話し言葉とWritingに用いるサインが融合しないと、日本語を表現するWritingにはならない。事実、日本語では漢字を導入した後からすぐ、万葉仮名のように日本語に合わせた音とlogogramの融合が図られる。
言語学では、この融合段階にRebusが重要な働きをしたと考えている。さて、Rebusとは何だろう? Rebusは判じ絵と訳されるが、文字やサインの読みから新しい意味を引き出すことを指している。ちょっとわかりにくいので、間違いなくひんしゅくを買いそうなダジャレを例に説明してみよう。例えば矢(や)と、缶(かん)を合わせると、ヤカンという新しい意味が生まれることがこれに当たる。この場合「やかん」は、「薬缶」でもいいし、「夜間」でもいいが、元の絵とは全く関係のない意味を持っていることが重要になる。

図3 Rebus 判じ絵 筆者が作成。
このダジャレのルール(Rebus原理)により、音とサインが結びつき、話される文章のどのような単語も文字化できるようになるのが、Writingの誕生だ。この過程についてPowellはそれまで使用されていたサインを使うだけのWriting(これをPowellはSemasiographyと呼んでいる)が「Words:単語」を使うWriting(これをLexigraphyと呼んでいる)に転換する過程だと述べている。
この考えをもう一度日本語を例に説明してみよう。日本語の話し言葉と、漢字はそもそも由来した場所が異なり、独立していると言える。最初漢字は、中国語として読まれ習われていたと思う。私たちが英語を習うようなものだ。ただ、日本語には中国語に匹敵するWritingがなかったため、漢字の一部を訓読みで道具として使い始めたのだろう。ただ、名前や、漢字にない単語、あるいは助詞など日本語本来の話し言葉の単語がないと、漢字だけで日本語の文章を文字化することが難しい。そこで万葉仮名がRebus原理で生まれ、それが表音文字(Phonogram)へと変化した仮名が生まれることで日本語の文章のWritingが完成した。これにより、人の名前を含め、日本語の話し言葉の全てが文字を用いてWritingすることができる。以上、日本語ではRebus原理はまさに万葉仮名として使われた後、仮名へと発展した。
実際、万葉仮名と同じような文字の使われ方が、メソポタミアやエジプトでも見られている。
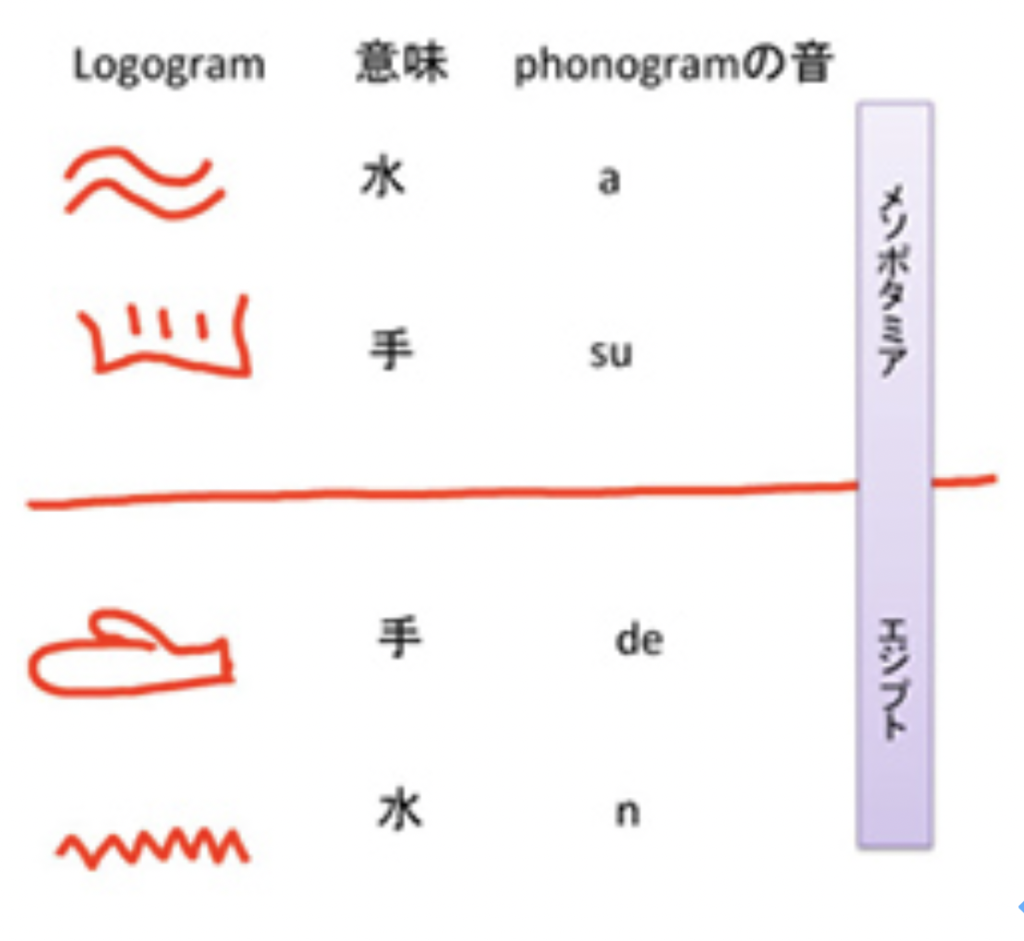
図4 メソポタミア及びエジプトでは絵文字の発音が、そのまま音を表す文字へと変化した。前述のPowellの著書を参考に筆者が作成
図4に示したのは、それまでlogogramとして使われていた、水や手といったサインが、万葉仮名のように、意味とは無関係に音の表現に変化した例を示している。
最後にもう一度強調したいのは、それぞれ独立していた話し言葉と、サインが Rebus 原理で融合し始めると、文字のボキャブラリーは急速に拡大、スピーチを全てWritingで表現できるようになる点だ。この相互作用の結果は、決して話し言葉が記録できるというようなレベルに止まらない。もともと、話し言葉は単語と単語の間の境界がわかりにくい。一方Writingでは明確に語と語の間に境をもうけることができる。両者が融合することで、言語自体の明瞭性は格段に上昇することになる。そして、持続性の高い文字が使えることで、ボキャブラリーの制限はなくなり、言語が無限に拡大を始める。このように、Writing と話し言葉が相互作用することで、片方ではなし得ない相乗作用を発揮し、言語の可能性が拡大することになる。
次回以降、このようなlogogramとphonogramの関係を、有名なシャンポリオンによるエジプト文字の解読や失読症を見ながらもう少し考えてみたい。
[ 西川 伸一 ]
2018年6月15日
これまでWritingという技術が誕生するまでの歴史を、表意文字と 表音文字が今も混在して、文字が進化した数千年の歴史をそのまま残しているWritingの生きた化石、日本語を参照しながら紹介してきた。もう一度強調したいのは、Writingが、音韻を基盤とするS言語(speech language:話し言葉を基盤とする言語)成立過程で形成された高次脳回路に、視覚を基盤とする新しいシンボルシステムが付け加わる形で統合して生まれた能力で、絵を描いて経験を表現することとは全く異なるという点だ。このため、中国発のWritingを用いる一部の言語を例外として、エジプトやメソポタミアのwritingは、カナのような子音と母音が組み合わさったシラブルを表現するsyllabogramを経て、子音と母音が完全に別れた完全な表音文字(アルファベット)によるWritingへと一直線に進化して行く。この過程については、これから解説するつもりでいるが、ただあまり先を急がずここで少し一息入れて、Writingにまつわる基礎的な問題をいくつか考えてみたい。
まず最初は、Writing/Readingの脳ネットワークについて考えてみたい。これを理解することで、なぜほとんどの言語でWritingがアルファベットに置き換わっていくかをある程度理解することができる。
これまで述べてきたように、言語を可能にする脳構造は、まず集団でゴールを共有して協力する社会を可能にする脳回路と、シンボルを用いたコミュニケーションを可能にする脳回路を基盤に生まれた原始言語の脳回路に始まる。その後この原始言語回路は、聴覚(音節)を媒体にするシンボル体系、そして視覚(文字)を媒体にするシンボル体系を順番に統合して、ほぼ全ての経験を表現して他の個体に伝達することができる無限の能力を獲得した。実際にはそれでもまだ完全ではなく、バーチャルメディアを媒体にした新しいシンボル体系を統合する過程が現在進行している。何れにせよ、最初の大きなステップは音節を媒体にしたシンボル体系が統合されたS言語の誕生と言っていい。そこでまず、S言語を可能にしている脳ネットワークを考えてみよう。
耳から聞いた音を言語として認識するシステムと、脳内で生まれた表象を喉頭、口腔、舌などをコントロールして発声する能力を基礎に、音韻を用いる複雑な音節を作ってシンボルとして使えるようになったのが、S言語と言えるが、この脳回路を図1のようにまとめることができる。
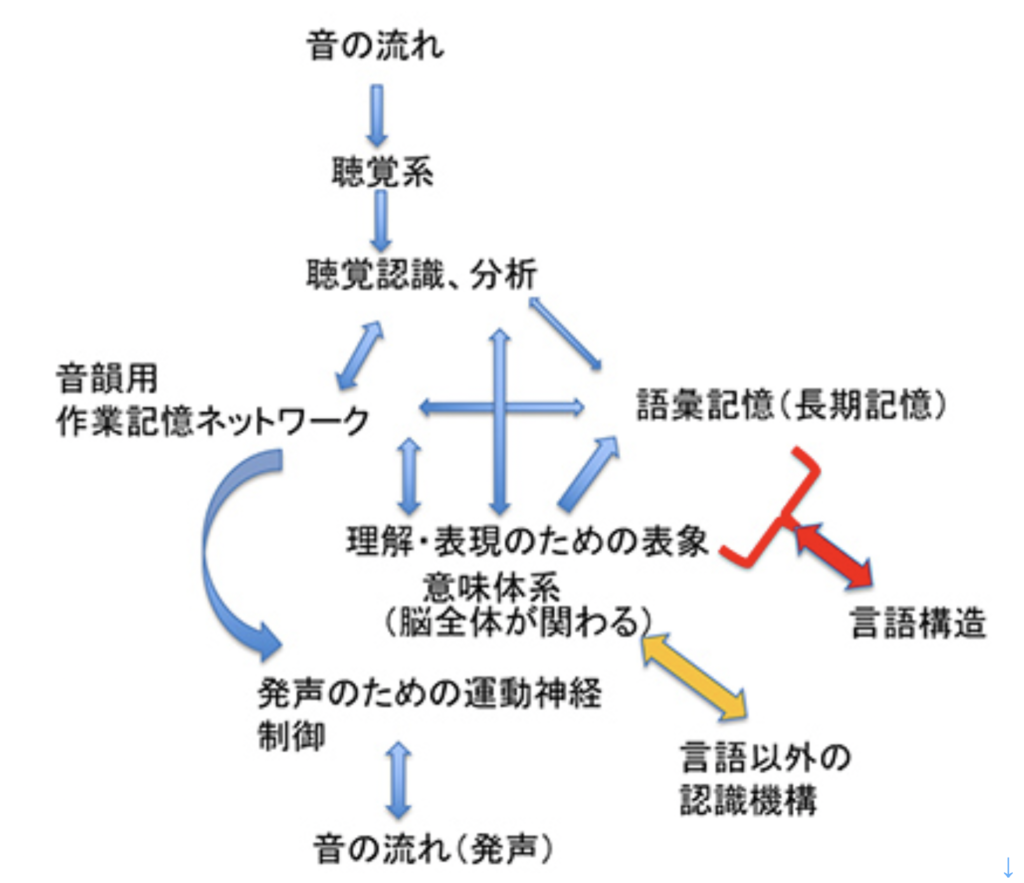
図1:S言語の脳回路。説明は本文参照。
この図を説明すると次のようになる。まず誰かが発する音を耳で聞く。突然人間の声が聞こえることもあるが、ほとんどの場合聞こえる以前から状況が判断され、聞こえて来るのが言語かどうか判断する準備が行われる。この過程で、言語とは別の社会性や、コミュニケーションを必要とする状況を判断する高次な意味体系が動員され、言語のコアネットワークにつながる。その後、聞こえてくる音節は、脳内に形成された語彙記憶だけでなく、すでに聞いたセンテンスや状況(他のインプット)から想像できるメッセージの意味と持続的に参照され、最終的に解読される。
このように聞こえた音節の塊は、多くの表象と参照されることで、メッセージの内容が解読されて行くが、この時うまく理解ができないと、以前紹介したような脳波上のN400が発生する(http://www.brh.co.jp/communication/shinka/2018/post_000006.html)。すなわち、把握された音節は、語彙の記憶だけでなく、常にメッセージ全体が持つ意味と参照され、解読されていく。この時重要な役割をもつのが、様々な脳内の表象を一時的に集めて参照するために必要な、短期の記憶に基づく統合を行う領域で、この図では音韻用作業記憶ネットワークと表現した。分かりやすくするため長ったらしい名前をつけたが、脳科学の人は、phonological buffer(音韻バッファー)と呼んでいる。
同じphonological bufferはもちろん、発語にも作業記憶ネットワークを提供する。発語の場合、経験したこと、記憶にあることなどまず伝えるメッセージが表象される必要がある。これも、最初からメッセージの詳細が全て決まっているわけではない。大まかに脳内で把握されている表象が、phonological bufferにインプットされ、音韻の流れとして表象されると、それに基づいて発語に関わる運動神経がコントロールされることで、文章が語られる。こうして生成される音韻は、また聴覚を通してメッセージの表象にフィードバックされ、思った内容が伝えられるよう、訂正が行われる。発生に関わる運動神経も、他の筋肉をコントロールするシステムと比べると、圧倒的に精緻なコントロールが行える複雑なものだ。徐々に進化してきたものだと思うが、声楽を聞くたびに大変なシステムだと感心する。
このようにS言語成立過程は、それ以前の言語のコア構造に対応する音韻のシステムを構築して、新しい言語の脳構造を作る過程としてみることができる。同じように、Writingの成立もそれ以前に完成した言語システムと無関係に成立することはなく、それまでに完成した言語構造の上に視覚認識を基盤にした新たなシンボルシステムを構築する過程と言える。言い換えると、既に成立していたS言語を可能にする言語の脳構造に寄生している。従って、WritingがそれまでのS言語と独立して成立することはない。逆に、S言語とほぼ同時期に見られたショーべやアルタミラの精巧な壁画は、言語のコア構造とはほぼ独立して存在していたと考えられ、その意味でWritingの起源にはなり得ないことも納得できるはずだ。
Writingが、S言語では難しい長期間の記録を可能にする為に発達したと考えると、自分の所有物を覚えておくために使われたトークンは、S言語と視覚認識を通した新しいシンボルとを連合させることに成功し、言語の脳構造の拡大に大きく寄与したと考えられる。この意味で、トークンはそれが表現するアイテムに直接対応するシンボルに一見思えるかもしれないが、実際には音節で表されるS言語の単語に対応するシンボルが誕生したと考えた方がいい。すなわち、WritingはS言語の拡大として、S言語の音節シンボルと常に対応をつけながら発展したと考えられる。その結果、図1に示した言語の脳構造は、図2に示すさらに複雑な構造へと発展してきた。
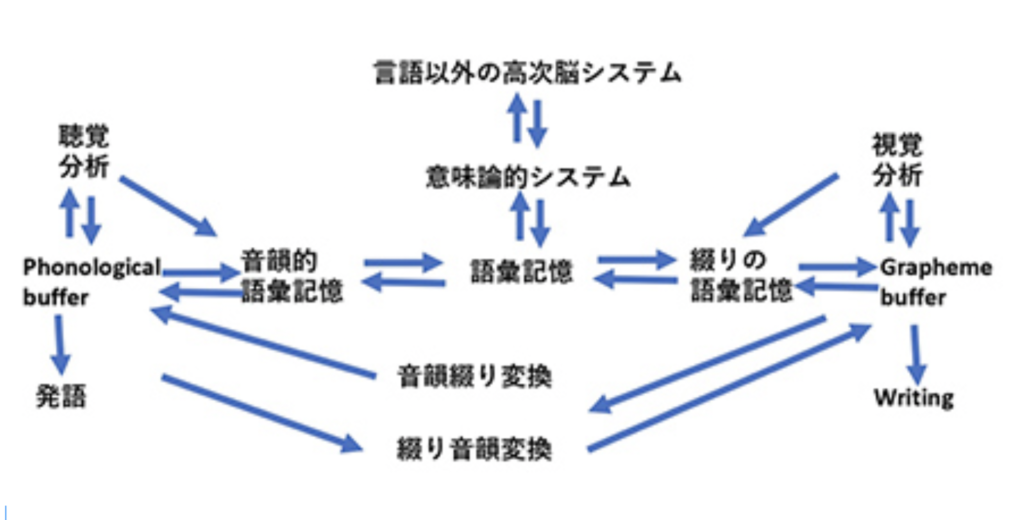
図2:話し言葉とWritingが統合された脳の言語構造。この図はDöhla
and Heim : Frontiers in Psychology 6: doi:
10.3389/fpsyg.2015.02045 を参考にして筆者が作成している。ただ、語彙記憶については変更している。
以前、言語を支配について発語を支配するブローカ野と言語の理解を支配するウェルニッケ野が関わると言う、極めて単純化した図式を紹介したと思うが(http://www.brh.co.jp/communication/shinka/2017/post_000012.html)、このように言語に関わる脳内ネットワークの条件を考えてくると、決してこんな単純な話でないことがわかる。事実、図に示した脳回路は、精一杯単純化した回路で、実際はこれの何千倍も複雑な回路が働いていると考えられる。このことは失語の患者さんの脳障害を高い精度で特定し、また機能的MRIや脳内各領域の結合がわかる拡散テンソル法などのイメージング(図3)による分析により確認されている。
NordicNeuroLab – Company Presentation from NordicNeuroLab on Vimeo.
図3言語により活性化される脳を機能的MRIと拡散テンソル法で分析したVimeocomにアップロードされたNordicNeuroLabによるビデオ
考えてみれば、発語に関わる複雑な運動調節を小脳の関与なしに行うことは出来ないし、作業記憶を海馬の助けなしに維持することは出来ない。事実、海馬の障害は、進行性の失語では最も可能性の高い原因になっている。
要約すると、言語は脳回路がネットワークとして組織化されて可能になる機能で、聞く、話す、読む、書くなど行動は異なっても、同じ脳回路がオーバーラップして関わっている。例えば書きながら読むと、外国語の単語も憶えやすいという経験は、異なる言語機能にも共通の脳回路が関わっている事を示す良い例だ。また、失語症で傷害される部位と症状を対応させる研究からも、同じ回路を異なる機能がオーバーラップして使っていることがよく分かる。現在失語症は、言葉を話すのが流ちょうかどうか、言葉の理解が出来ているかどうか、そして他人の言葉を復唱できるかどうかの3種類の症状の有無で8種類の類型に分類している。例えば「流ちょうに話すのだが、内容に論理性がなく、ほかの人の言葉を理解しておらず、他人の言葉を復唱も出来ない」と、ウェルニッケ失語と診断され、「他人の話を理解できるのだが、言葉がたどたどしく、復唱ができない」場合はブローカ失語と診断される。こうして分類されたそれぞれのタイプの失語では、音韻によるS原語とともにWrintingも同様に侵されることが多いが、これも共通の回路を示す良い例だ。
しかし、それぞれの機能に比較的特異的に関わる回路も存在する。この結果、上の8分類だけでなく、それぞれの機能に特化した症状も現れる場合がある。 その典型が、失読症(dyslexia)や失書症(agraphia)で、きわめて多彩な症例がこれまで報告されている。
そこで最後に三井記念病院から1997年にNeurologyに報告された日本人Writin特異的障害を持った2人の例を紹介して終わろう(図4)。

図4 1997年に三井記念病院からNeurologyに発表された、仮名や漢字のwritingが選択的に傷害された患者さんの報告。
第一の症例は76歳の女性で、専門医を訪れたときは、軽い非流調性失語と、計算が出来ないという症状があるが、理解や復唱、聞いたり読んだりするのはほぼ正常だった。ところが、文字、特に漢字を書くのが強く傷害され、漢字特異的な失書症と診断された。異常の原因は脳梗塞で、中前頭回とそれに隣接する中心前回に傷害が見られた。
第2例目は69歳男性で、流ちょう型の失語と診断できるが、理解や、復唱には異常は認めていない。流暢に話が出来き、聞く理解も正常であるにも関わらず、仮名を書く能力が強く低下している一方、漢字の読み書きは比較的保たれていた。第一例と同じで、計算能力の低下が見られる。原因は同じく脳梗塞で中前頭回が傷害されていた。
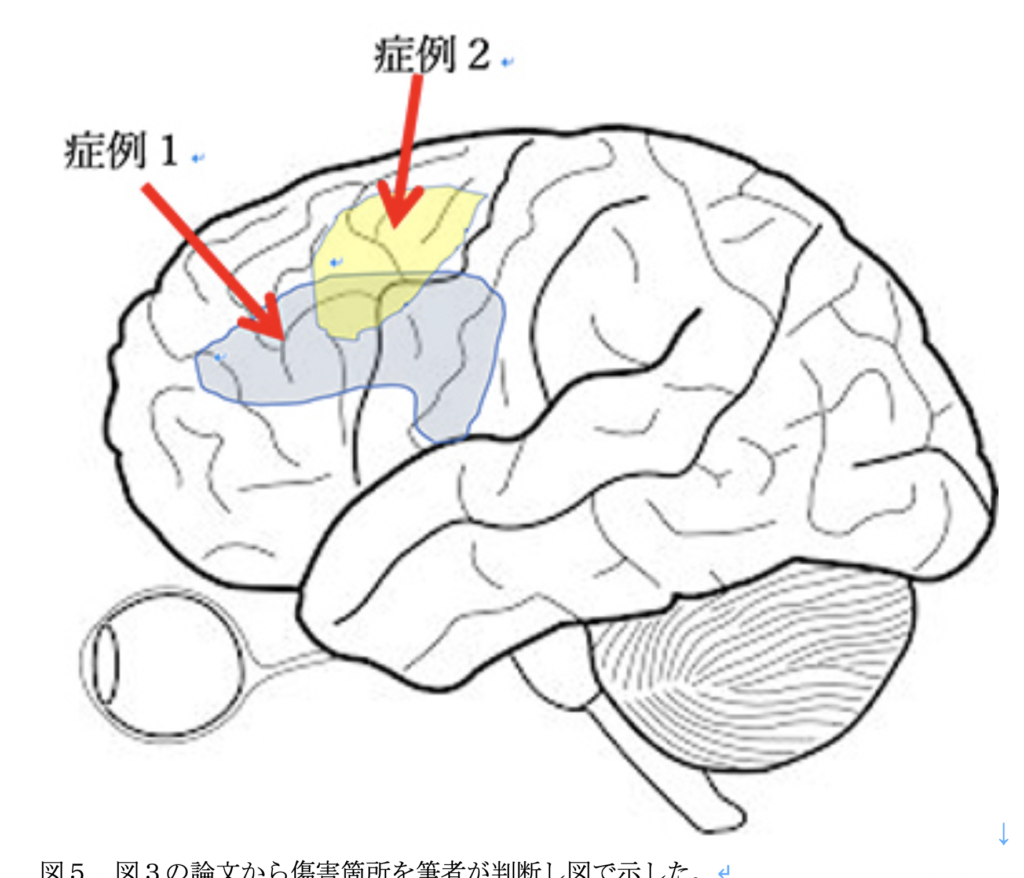
図5 図3の論文から傷害箇所を筆者が判断し図で示した。
病変もオーバーラップしているのに、少しずれると、仮名と漢字が別々に障害されるのを見ると、失語を脳回路から理解することの難しさがよく分かる。裏返せば、何度も繰り返してきたように言語の脳回路の複雑性を示している。
以上今回の話はつぎのようにまとめられるだろう。
言語の脳回路は、原始的構造を拡張しながら現在の形に到達した。則ち、原始言語構造にS言語を可能にするネットワークが加わり、そこに新たにWritingを可能にしたネットワークが統合されている。このことは、後から統合されたネットワークは、それ以前のネットワークの制約があるため、全く独立しては存在し得ないことを意味している。この結果、例えばWritingに関わる失読や失書にはS言語の失語も多かれ少なかれ合併する事になる。
しかし一般的に失読症と言うときは、ほかの症状が合併しない、例えばダビンチやアインシュタインなど天才が含まれる状態を指すことが多い。次回は、この純粋な失読症について見て行く予定だ。
[ 西川 伸一 ]
2018年7月2日
前回紹介したように、hearing/speaking(以後話し言葉)とreading/writing(以後読み書き) は互いに言語に関わる様々な脳回路を使っているため、例えば脳卒中や外傷で脳組織が障害されて失語症が起こる殆どの場合、どちらも影響され、どちらかが独立しておこることは稀だ。
しかし、これとは別に、明確な脳の障害はないにもかかわらず、発達期に読み書きが強く障害される人たちが存在し、このようなケースを特別にディスレキシア(失読症)と呼んでいる。このような失読症は、「知能は正常で、別段神経的異常がないにも関わらず、読むのが不正確で、時間がかかる」症状を示す状態、と定義出来る。
ただ、話せるようになっていても読み書きが難しいのは誰もが経験してきたことだ。言葉を理解し、話すことが出来ることが読み書きの必要条件だが、話せるからといって読み書きが出来るというわけではない。読み書きが出来るためには、話し言葉の言語脳のまわりに、各人が多大な努力を払って読み書きの脳を形成し統合させる必要がある。よく考えてみると、識字率が問題になるように、ちょっと前まで世界中の言葉を話す人間のほんの一部しか読み書きが出来なかった。明治維新に自分の名前を書ける鹿児島の女性は10%に満たなかったと言われている。これは、殆どの人が読み書きの教育を受けていなかったからだ。Writingが数千年の歴史を持つといっても、その恩恵にあずかっていたのは、新たに読み書きの教育を受けることが出来る、ほんの一握りの人たちだったのだ。このことは、ほかの人間とコミュニケーションを繰り返すうちに自然に学習する話し言葉と異なり、読み書きの習得は、系統的な「教育学習」が必要な、個人の成長にとって大事業であることを示唆している。
私達が話し言葉をマスターするのは大体3歳を過ぎた頃だが、親が話しかけるなどある程度の努力はしたとしても、話すための系統的教育を行うわけではない。ところが3歳をこえて話し言葉をマスターする時期になると、今の親は読み書きを教えるための様々なプログラムの宣伝に目を奪われるはずだ。このような宣伝は、「本当にすぐ読み書きを教えないと、私達の子供の発達は遅れるのか?」と多くのお母さんを心配させ、混乱に陥れている。しかし本当は、いつから読み書きを教えればいいのか? 早いほど良いのか?については、今も論争が続いている。恐らくこの問題に科学的決着をつけるためには、長期のよくデザインされたコホート研究が必要になる。ただ、これは簡単ではない。強制的に早い段階から読み書きを教える事により、取り返しがつかない結果を招く可能性すら存在する。次回議論するように、我々が異常と判断した性質が、本当は新しいイノベーションを生み出すのかもしれない。結局、多様な個性を大切にすることが最も重要なことで、子供の習いたいという欲求と、それに答える系統的プログラムを用意しておくのが最も重要だろう。
繰り返すと、この系統的な教育が必要であると言う点が、話しことばと読み書きの最も重要な違いで、あらゆる言語、あらゆる時代で共通だ。この過程を通過しないと、文盲で終わる。1つの例を見てみよう。今から3千年ほど前、シュメールの楔形文字を習うため、おそらくエリート階級から7歳前後の子供たちが選ばれ、何年もかけて読み書きを習ったe-dubbaと呼ばれる学校があったことが知られている。そこで学生が手習いに使ったタブレットが残されているが(図1)、先生の手本に従ってビッシリと手習いが行われており、系統的に読み書きが教えられたことが分かっている。教育は厳しかったようで、教師に鞭で打たれたことが書き込まれたタブレットもあるらしい。

図1 シュメール楔形文字の練習に使われたタブレットのかけら
(出典:Wikipedia)
このように考えると、ディスレキシアとは、自然に獲得されたS言語に教育を通じて読み書きの脳を統合する過程の異常と定義する事が出来る。このメカニズムについてこれまで様々な説が出されているが、個人的には自閉症などと同じで、多様な状態の集まりだと考えている。ただどの仮説を支持するにせよ、一致しているのは、ディスレキシアには高い遺伝性がある点だ。実際、ディスレキシアの子供の親族には、探せばだれか読み書きが遅かった親族がいる場合が多い。また子供がディスレキシアと診断され、自分もディスレキシアだったことを親が認識することも多い。
遺伝性を調べるために行われた大規模な双生児研究によると、ディスレキシアが両方で一致する率は、一卵性双生児で68%、二卵性双生児で38%に達する。またゲノム解析により相関が認められた遺伝子座が10箇所以上明らかにされている(図2)。しかし特定された領域同士の関係は現在も解析が進んでおらず、ディスレキシアを分子ネットワークの言葉で説明するには、まだまだ時間がかかると思われる。

図2 これまでディスレキシアと相関が認められた遺伝子座 Peterson and
Pennington Annual Review of Clinical Psychology 11:283の図を改変して用いている。
何れにせよ、遺伝性が高いと言うことはディスレキシアという状態が、読み書きを習う以前から存在することを示唆している。この可能性を確かめようと、家族歴からディスレキシア発症リスクの高い子供の脳波やMRI検査を幼児期に行い、その後読み書きを学習する時期まで追跡するコホート研究が行われた。2008年にフィンランドから発表された論文では、外れた調子の音が急に現れたときの反応が、最終的にディスレキシアを発症した新生児では明らかに低下していることを報告している。また、ディスレキシアは発症しなかったが、家族歴から遺伝的リスクが高いと診断された子供も、軽度だが反応が低下していることも示している( Leppänen et al Cortex 46:1362,2010)
脳領域についての詳細は省くが、読み書きを習う前からディスレキシアの発症リスクをMRIによって予知することが出来るという論文も最近発表された(Clark et al, Brain 137:3136, 2014, Raschle et al, Neuroimage 57:742, 2011)。これらの結果は、ディスレキシアの多くが、遺伝的な変異によりおこる脳構造の変化により生まれつき決まっていることを示唆している。
では、ディスレキシアの背景には、どのような脳機能の変化が存在するのだろうか?この問題に関しては、従来から2つの考え方がある。1つは先に述べたフィンランドの研究から分かるように、連続する音や視覚刺激に対する反応の異常によって読み書きが阻害されるとする考え方だ。もう一つは、話しことばの脳が形成されるとき、先天的・後天的にすこし違った構造が出来てしまい、これが新たな読み書き脳の統合を阻むという考え方だ。これはPhonological Deficit仮説と呼ばれており、ディスレキシア理解の現在の主流になっている考え方だ。
個人的にはディスレキシアは多様な脳の状態を反映しており、どちらか一方で決まりという話ではないと思っている。例えば2013年、私のブログで紹介したが、ハーバード大学のグループは文章を読むのが苦手な子供に、活字が詰まった本ではなく、スマートフォンの画面に少しづつ文字が現れるようにすると、一般児と同じレベルで文章を理解できるようになった事をPlosOneに発表した(http://aasj.jp/news/watch/509)。おそらくディスレキシアと診断される子供の中には、このように多くの文字に一度に晒されると気が散って集中できないケースもあると思う。自分自身の体験から考えると、日本語を読むときはあまり感じないが、英語になると電子媒体で読む方が頭に入ってくる。私の場合せっかちで気がせくため、多くの行に目が飛んで集中できなくなることが英語を読むのが苦手な原因になっているが、同じ様な子供は多くいるのではと思っている。いずれにせよ、どんな小さな現象でも、私達がどう文字を解読しているのか考える上で役に立つ。
繰り返すが、私達の言語脳は読み書きの脳が統合されることで質的変化を遂げる(例えば時間や場所を越えた間接的なコミュニケーションが可能になる。)読み書き能力の習得は私達が一生で経験する学習の中でも最も大事業だ。すなわち自然に形成された話しことばによる言語脳を、読み書きを能力が統合した質的に新しい脳へ学習を通して発展させる事業だ。その意味で、他の学習とは複雑度が格段に違うと予測できる。ただ、この事業は自然に習得した話しことばを土代に行われることから、ディスレキシアはこの土台の異常だとする考えが存在し、Phonological deficit仮説と呼ばれている。
次回は、ディスレキシアの脳について考えた後、ディスレキシアの人たちにはなぜ天才と言われる人が多いのかを考える。
[ 西川 伸一 ]
2018年7月17日
ディスレキシアは「読み書き」という視覚に関わる言語の遅れが主症状として目立つため、最初視覚システム特異的な発達障害ではないかと考えられていた。実際、前回紹介した本を読むのが苦手な子供に、スマートフォンで、少ない文字数を大きな文字で提示することで、文章の読解が正常になるという症例は読むことに特化した、視覚依存性のディスレキシアの存在を示している。
このような方向の延長で、ディスレキシアが視覚システム主体の障害であることを最も強く主張するのがMagnocellular deficit仮説で、ディスレキシアの原因が、網膜神経節のパラソル細胞と、そこから神経投射を受ける外側膝状体の中のMagnocellular層を含む視覚回路の障害だと考える仮説だ。
光に反応する視細胞の興奮はまず双極細胞やアマクリン細胞などの中間神経に集められる。これに続いて、中間神経に集まった網膜情報を、視床を中心に視覚に関わる各脳領域へ伝達するのが網膜神経節細胞で、大型のパラソル細胞と小形のミジェット細胞に分かれる(図1)。
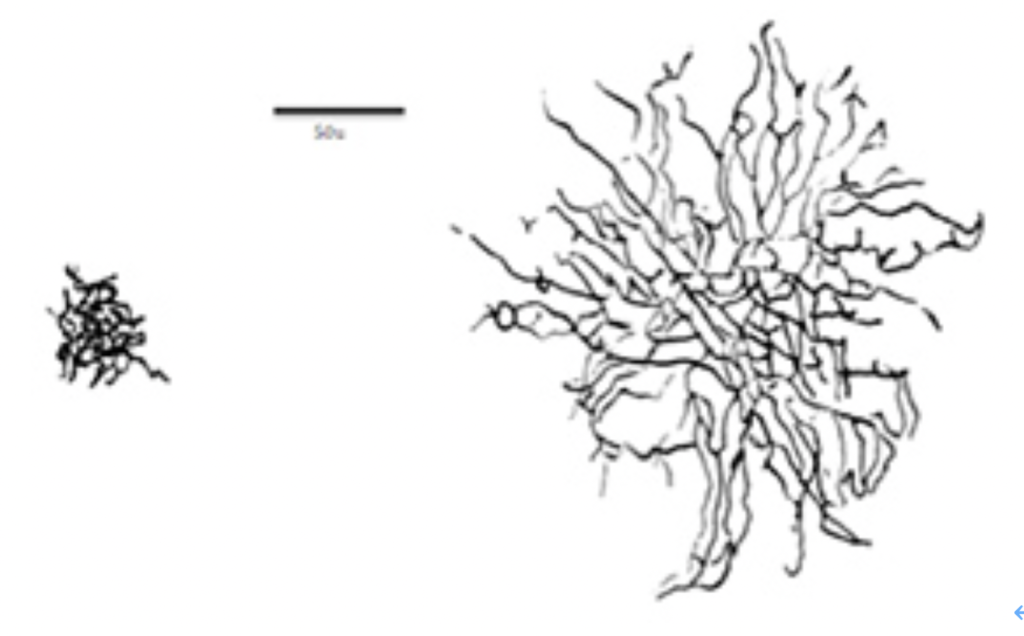
図1 網膜神経節のパラソル細胞とミジェット細胞。
出典:Wikipedia
このパラソル細胞は形からわかるように多くの中間細胞と結合しているため、解像度の高い視覚認識は全てミジェット細胞にまかせている。代わりに、弱い光に素早く反応出来るので、動く物を追ったり、あるいは文字をたどる時の時間的変化を把握してタイミングをとる過程に関わっている。このパラソル細胞からの神経投射は外側膝状体のMagnocellular layerとよばれるやはり大型の神経細胞に投射し、光の変化をいち早く捉え、他の視覚シグナルに情報を提供する回路を形成している。
くり返すが、この回路は解像度の高い視覚には関わらないため、文字の認識の際も文字の形の詳細の認識とは無関係だ。ところが、ディスレキシアの子供ではこの回路が機能的に障害されているとする研究結果が最近集ってきた(例: Pammer and Wheatley, Isolating the M-cell response in dyslexia using the apatial frequency doubling illusion, Vision Research 41:2139, 2001)。更に、剖検例の解剖学的解析や、MRIを用いた画像解析からも外側膝状体のMagnoccelular areaの発達が遅れていることが指摘され(Müller-Axt and von Krigstein, Altered structural connectivity of the left visual, Current Biology 27:3692, 2017)、少なくとも一部のディスレキシアでは網膜神経節から外側膝状体、そして皮質の視覚野への回路異常が基盤にあることは広く認められるようになった。何よりも、薄明かりの中で動体を追いかける訓練などでMagnocellular pathwayの回路を発達させることで、ディスレキシアが改善されることが明らかになり、Magnocellular hypothesisはディスレキシアを視覚回路の異常として捉える最も重要な考えとして定着した。
一方、多くのディスレキシアの子供たちが、学校で文字を習い始める前から、話しことばの発達が遅れたり、長い単語を正確に発音できず、アルファベットがごちゃ混ぜになったり(例えば「Teddy bear」を「beddy tear 」)、言いたいことがうまく言えなかったり、韻をふんでいる文章が苦手だったりと、読み書きだけでなく、話しことばの発達が遅れていることが認識されていた。このことから、ディスレキシアは純粋に読み書きという視覚に特異的な障害ではなく、言語脳全体の発達障害ではないかという考えがPhonological deficit仮説だ。
この仮説は、文字を習う以前、私達が話し言葉を学習する過程が、質的に異なる2段階に分かれることを前提として考えられている。最初の段階は聞き覚えた音を自動的に並べただけのコミュニケーションで、この段階の言葉は本能的で自分の話している音を正確に認識しているわけではない。従って本人の頭の中でTeddy bear が表象されていても、発音している単語を意識的に把握していないためbeddy tearと言ってしまうことになる。おなじ事は単語だけでなく、おそらく文法的にも言えると思う。普遍文法といえる自動的、本能的な原始的文法は、最初の段階の特徴だ。ところが文法的にも正確な言葉の並びを使うようになる次の段階には、質的な能力の転換が必要になる。おそらくこの転換の中には、流暢な言語を話すために必須の、様々な表象を一時的に保持しておく作業記憶が言語に対応することが必要で、最初の段階ではまだ発達し切れていないと考えている(個人的意見)。
ところが読み書きを習う前には、作業記憶がしっかりと言語に対応出来るように準備され、聞いた言葉や話そうとする言葉を作業記憶領域に保持し、それを音節レベルでハッキリと意識できるようになる。これがphonological awarenessと呼ばれている能力で、自分が話している言葉が特定の音の並びで出来ていることを意識するためには欠かせない重要な能力だ。子供たちのphonological awarenessを可能にする言語回路の形成が遅れると、話し言葉だけではなく、当然視覚を通して認識する書かれた文字と話しことばを対応させる過程にも遅れが生じ、ディスレキシアとして現れると考えるのがphonological deficit 仮説だ。この考えは、子供の言語能力や行動学的機能検査だけから現象論的に把握されるだけではない。最近では、脳波計や、MRIを用いた画像研究からもこの考えを支持する結果が出されている。
例えば、生理学研究所(愛知県岡崎市)の定藤らのグループは、我が国のディスレキシアの子供が仮名を使うときの問題を機能的、脳画像的に調べ、読み書きだけではなくphonological awarenessの発達が遅れると共に、一般児でphonological awarenessの発達により反応が高まる紡錘状回の反応が、ディスレキシア児では低下していることを明らかにしている(Koeda et al, Dyslexia: advances in clinical and imaging studies, Brain and Development, 33:268, 2011)。この結果は、英語圏のディスレキシアの子供と臨床的にも、画像的にもほぼ同じ結果で、表意文字とともに音節表記文字(完全な表音文字であるアルファベットとは異なる)の仮名を用いる我が国の子供のディスレキシアも、基本的にはphonological deficitを基盤に出来ていることを示している。中国語ではどうなのかまだ把握していないが、画像レベルで客観的な変化が明らかになることで、ディスレキシアの表現の言語による違いのみならず、言語そのものの構造の新しい側面が見えてくるのかもしれない。以上の結果を図2のようにまとめてみたので、何かの参考にしてほしい。
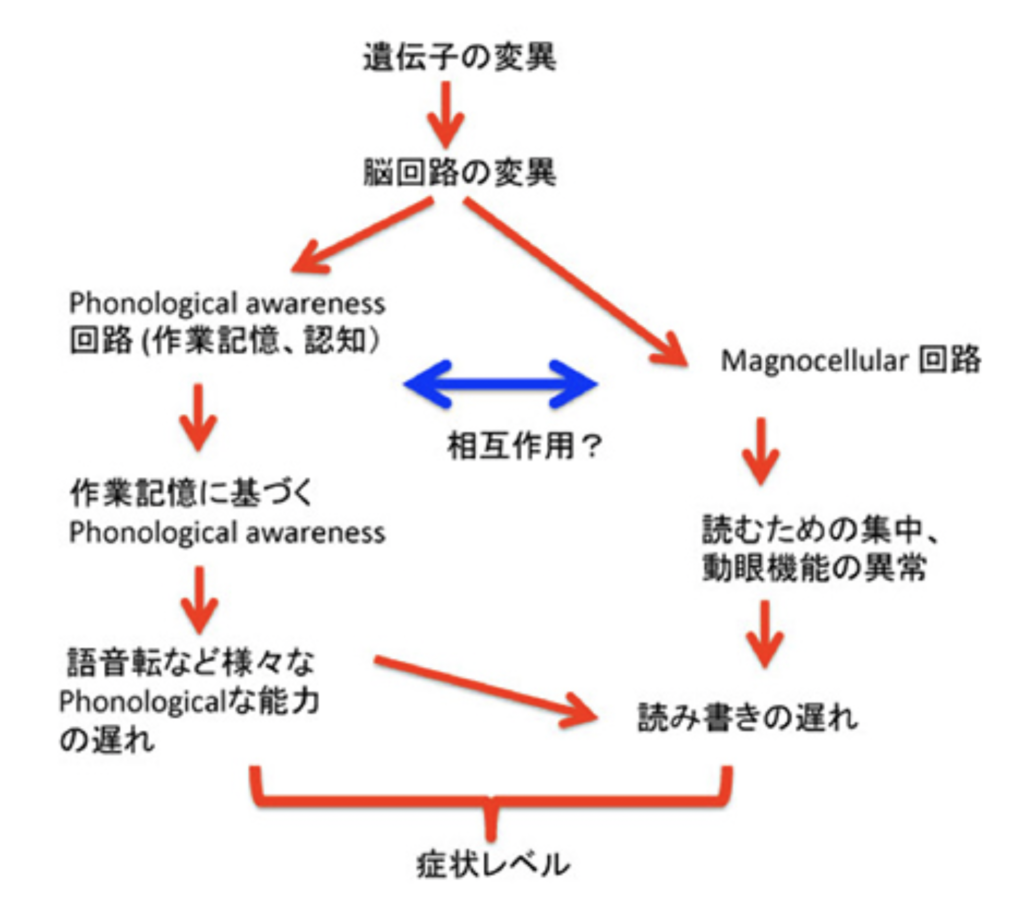
図2 ディスレキシアに関する2つの仮説。どちらの仮説も、遺伝的な脳回路の変異が、能力や行動の異常に繋がると考えている。
これまでディスレキシアを発達障害として扱ってきたが、ディスレキシアの本当のおもしろさは、子供時代にディスレキシアを経験した人たちの中に、大人になってから類い希なる能力を発揮する人たちが多い事が知られている点だ。例えば図3に示したBrock Eide(ブロック・アイド)著『The dyslexic advantage』はこのことを一班の人に伝えようと書かれた本だ。読んでみると、ディスレキシアについての本と言うより、ディスレキシアを持つという事の素晴らしさについて書いた本だ。

図3:ディスレキシアの脳の優位性を強調する本。
内容は、ディスレキシアの著名人をリストし、それぞれの才能から、ディスレキシア脳が優れている理由を解説している。
出典:Amazon.co.jp
ディスレキシアの著名人には、古いところでダビンチ、最近ではアインシュタインやエジソンが良く知られているが、この本ではアメリカ人なら知っている様々な名前の長いリストが示されている。そして、これら著名人の成功は、ディスレキシアの脳が、
を持っているからだと強調している。
著者はお医者さんらしいが、内容を詳しく読むと論理が強引で、科学的か?と問われると私は否定的だ。例えばディスレキシアの脳がもつ優れた空間イメージについては、まずノーベル賞に輝いたグリッド細胞の話しが引用され、この機能とディスレキシアの関係を、空間イメージを住宅ディベロッパーとしての仕事に生かして大成功した、ディスレキシアの富豪の話しで裏付ける、といった強引な論理構成が随所に見られる。その意味でお勧めの本と言うわけではないが、ディスレキシアを発達障害として否定的にとらえることの間違いを正す意味では評価できると思う。
この考え方の背景には、読み書きを習うことで、私たちが犠牲にせざるを得ない能力があるという考えがある。例えば、視覚的経験の詳細を決して忘れないサバン症候群の人の特殊能力を知ると、どうしてこんな事が人間の脳に可能かと驚くが、逆に一般人なら普通に持っている様々な能力がサバン症候群の人では失われていることも確かだ。
多いか少ないかは別にして、ディスレキシアの中に、アインシュタインやエジソンがいることを知ると、逆に読み書きを習う過程で私たちの脳が失う能力があることに気づく。そしてこの印象を科学的に裏付けられるのかについて、実際に多くの研究が行われている。ただ、最近の論文を見渡すと、様々なタイプが含まれるディスレキシア全体を対象として調べると、空間認識や処理能力に一般児と大きな違いはないとする論文が多い印象がある。
このことから、ディスレキアがあるから必ず空間認識が高まるわけではないが、きわめて優れた空間認識脳や抽象化能力をもつ人は、ディスレキシアの傾向が高い事については、十分あり得る話しだと思っていいだろう。例えばディスレキシアの人には鏡文字として知られる、右から左へとながれる文字を苦労なく書ける人たちがおり、ダビンチもその一人だが、(図4)この能力は文字を文字としてではなく、1つの自由な画像として捉えることが出来る能力を反映している様に思える。読み書きを習うことが、このような能力を犠牲にすることだとすると、読み書きを習う過程で、私達の脳をもう一度構成し直す大事業が進んでいることを意味する。しかし、この大事業のおかげで、phonological awarenessはより永続的な記録性を獲得し、人間社会を質的に変革する事が出来た。

図4 レオナルド・ダ・ヴィンチの鏡文字
出典:openculture.com
このあたりのことを議論し始めるときりがないので、最後に科学に基づいて読み書きの脳について最新の研究結果をわかりやすく紹介しながら、ディスレキシアについても述べた本を一冊紹介して終わろう。本は『プルーストとイカ』という面白いタイトルがついているが、副題が「読書は脳をどのようにかえるか?」で、読み書きの歴史からディスレキシアまで、発表された論文に基づいて議論が行われている。今だから正直に告白するが、この本を読めば私がこれまでWritingについてくどくど書いてきたことを詠む必要はなくなる。是非一読してほしい。

図5:読み書きの学習と脳について書かれた素晴らしい本。
著者のMaryanne Wolf(マリアン・ウルフ)は現在タフツ大学教授で、ご子息がディスレキシアと診断されている。これまで紹介したWritingに関する本と比べると、より一般向けだが、深い科学知識に裏付けられている。
出典:Amazon.co.jp
[ 西川 伸一 ]
2018年9月3日
すでに、トークン、pictogram(絵文字)からlogogram(表意文字)と文字が進展する過程を見て来た。今回から文字によるWritingがsyllabogram (音節表記文字)、そしてphnogram(表音文字)へと進展する過程を、メソポタミアの楔形文字や、エジプトのヒエログラフを題材に見ていこうと思っている。重要なことは、これらのWritingは既に滅びた文字で、近代に入ってもう一度解読された点だ。そこで、本題に入る前に、古代文字がどのように解読されたのかを有名なシャンポリオンの例で見てみよう。ただこの分野についての私の知識は以前紹介したWritingについて書かれた何冊かの本(例えばBary Powellの著書:http://www.brh.co.jp/communication/shinka/2018/post_000010.html)や、図1に示すようなヒエログリフの読み方について一般向けに解説した本から仕入れていることを断っておく。

図1 NYロポリタン美術館で買い求めた古代文字読解のための解説書。
さて、エジプトで文字が現れるのは紀元前3000年前頃と考えられるが、様々な変遷を遂げながらも紀元400年までは用いられていた証拠が残っている。ただ、エジプトが最初はローマ帝国に、そしてビザンチン帝国、最後にイスラム帝国に支配される過程で、ヒエログリフは文化のあらゆる場面から消失してしまう。このため、エジプトの遺跡に残るヒエログリフは新たに解読しないと理解することは出来なかった。
このヒエログリフを初めて解読し、新しくエジプト学を確立した立役者が、皆さんもご存じのジャン・フランソワ・シャンポリオンで、最も古い絵文字や表意文字についてはまだわからない事も多いが、エジプト王朝の多くの記録についてはかなり理解できているといっていいだろう。

図2 ロゼッタストーンのヒエログリフをてがかりに、ヒエログリフ解読に成功したシャンポリオン。
Bary Powellの「Writing」によると、シャンポリオンは生まれながらに言語学の天才だったようだ。子供の頃からギリシャ語やラテン語をマスターし、なんと16歳でエジプトの地名がコプト語起源である点についての論文まで発表していたようだ。パリではペルシャ語、アラビア語を研究し、19歳でグルノーブル大学の教授の席を射止めている。
この天才をエジプト文字解読へ駆り立てたのは、1822年に送られてきたアスワン近くのオベリスクに書かれた文字の写本だった。このオベリスクは紀元前2世紀にプトレマイオス8世と、その妻クレオパトラ三世により建立され、ヒエログリフと共にギリシャ語で両者の名前が刻まれていた。
おなじ様に、ギリシャ語とエジプト文字が書かれているエジプトの遺物で最も有名なのは、現在大英博物館の人気展示物ロゼッタストーンだが、ここにもギリシャ語でプトレマイオスの文字とそれに対応するヒエログリフが存在している。シャンポリオンはヒエログリフの中で音を受け持つ表音文字を解読するには固有名詞がキーになると考えて、この3つの名前が書かれたヒエログリフ(図3)から解読を始めている。
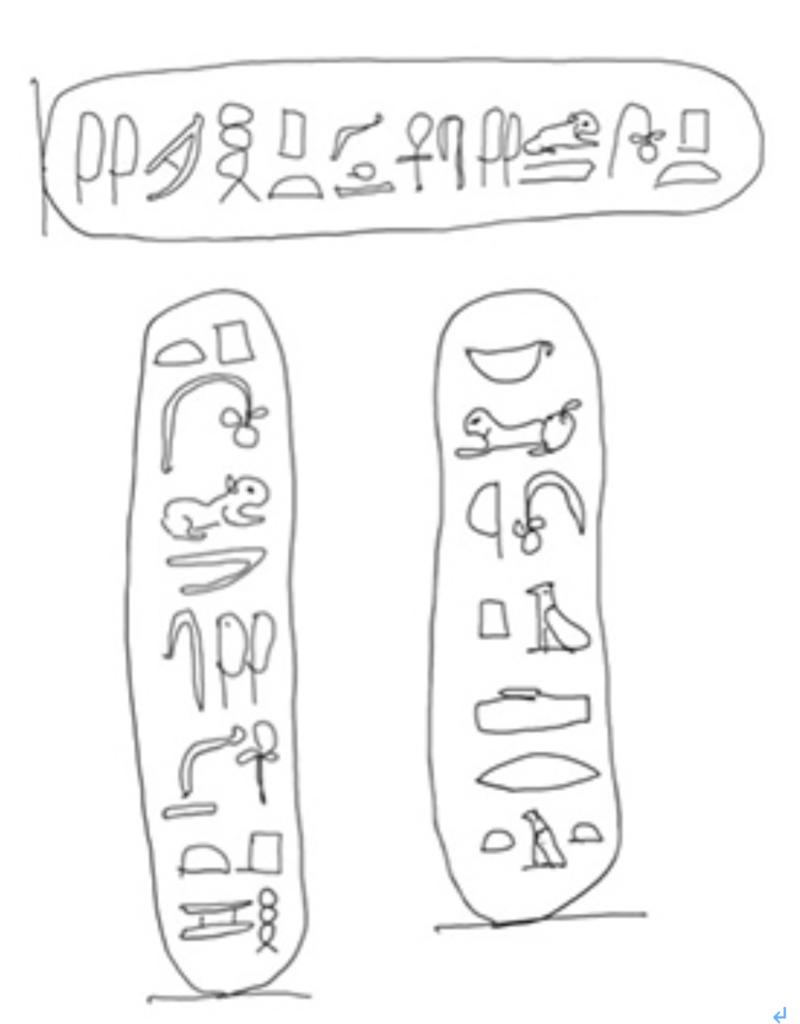
図3 シャンポリオンが解読に使った3つの名前。
全て私の手で書き写しており、正確ではないが、一番上の横書きがロゼッタストーン由来。下の2つの縦書きがBankes Obeliskに書かれた文字。それぞれ、プトレマイオス、プトレマイオス、クレオパトラに相当する
これらのサインが名前に相当することは、文字が全てカートリッジに囲まれていることから区別できる。また、これらの文字がプトレマイオスやクレオパトラであることは、ギリシャ語で書かれた文章から推察できるが、残りの文字がどう対応しているのかは、全くわかっていなかった。名前の場合は表意文字では表せないため、当時から表音文字が使われていたと推察できるので、この2つの名前が共通に持っているP(ピー)の音に対応する文字を探すと、プトレマイオスでは最初に、クレオパトラでは4行目に来る

がこれに当たる。同じ様に、両方に存在する
は、共にOの音に相当し、ライオンは
L、
はE,

ではTなど、他にもM,S,R,A などが解読出来る。また同じTをクレオパトラでは

で表現していることもわかった。
このように、カートリッジに囲まれた名前をさらに集めて、シャンポリオンは最終的に40の表音文字を特定することに成功している(図4にエジプトの表音文字リストを示す)。

図4:ヒエログリフの中の表音文字。(Wikimediaより:https://commons.wikimedia.org/wiki/File:Hieroglyph_picture_write_alphabet.jpg)
ただ、ヒエログリフをアルファベットと考えるのは間違っている。例えばロゼッタストーンに書かれたギリシャ語は約500語存在するが、同じ内容が表現されているヒエログリフではなんと1419種類ものサインが使われている。このことを、今皆さんが読んでいる私の原稿のタイトル「表意文字から音節表記文字へ」を例に当てはめて考えると、このタイトルでは5つの単語のために、10の異なるサインをもちいている。このように音をあらわすsyllabogramの使用が始まると、文字数は増加する。
実際図4のアルファベットに対応する文字は、一見するだけで絵文字に近いことがわかる。事実最初の鷹はAlephと読むことができるが、表音文字としてはAの音として読まれる。これは、私達が漢字から表音文字としての万葉仮名を当てたのとおなじだ。また、上に示したアルファベットも、実際のアルファベットとは異なり、子音と母音が合わさったsyllabogram(音節表記文字)で、この点でも日本語の「か:Ka」「じ:Ji」などとおなじだ。実際には、ヒエログリフはさらに複雑で、より多くの音節が集まった文字も存在する。例えば

は表意文字としてはコガネムシだが、syllabogramとしてはhpr(kheper)と読まれる。
これらの他に、ヒエログリフでは続く単語のカテゴリーを示すためだけにサインが使われ、例えば

:太陽は時間を表していることを示す場合の接頭詞として使われたりしている。
さらに日本語のWritingと類似しているのは、その後、表意文字をそのまま表音文字に使い回していたヒエログリフが、さらに抽象化され、単純化されるときに宗教的儀式に用いられる神官文字と、一般に用いられるdemotic文字にわかれる。これも平仮名と、カタカナに分かれるのと似ているといえないだろうか

図5 ヒエログリフから生まれた2種類の文字。Wikimedia commonsより。
このように、時に絵文字、表意文字、音節表記文字がWritingにつかわれるというのは、我々が現在使っている日本語とそっくりで、逆にいうと日本語が、エジプト文字で言えば紀元前に相当するWritingを使っている、まさに奇跡のシステムであることがよく分かってもらったと思う。
次回は、Writingの最後、エジプトやメソポタミアの音節表意文字が、アルファベットに変化した過程を追いかけてみる。
[ 西川 伸一 ]
2018年9月18日
繰り返しになるが、日本語のWritingは、絵文字(pictogram)を下敷きに抽象化されて生まれた中国由来の表意文字(logogram)漢字をそのまま使い、この漢字から音だけを取り出すrebusという手法を用いた万葉仮名を表音節文字(syllabogram)としてまず開発したあと、そこからさらに我が国独自の2種類の音節文字、仮名を発展させ、最終的に漢字と仮名の両方を用いるlogosyllabaryと呼ばれるシステムを完成させた。そしてこのlogosyllabary体系は既に1000年以上使われている。
日本人にとってこの歴史は何の不思議もないことだが、他の言語の文字の歴史を考えると、logosyllabaryが使われていること自体、まさに奇跡と言っていいように思う。しかし、日本語がこのシステムを維持できたおかげで、今やアルファベットやアラビア数字など、便利と思える表記体系は全て統合できる。実際、表音節文字だけでなく、アルファベットのような完全な表音文字(phonogram)すら、外来語やローマ字として一般の日本人が使うようになってきた。まさに、あらゆるWritingを吸収できる融通無碍の奇跡の表記体系と言って良いだろう。これに対し、例えば英語を考えてみると、英語を話す人たちに寿司を食べることがポピュラーになったとしても、「寿司」という文字は使われず「Sushi」と書かれる。今後日本語がどう発展していくのか、興味は尽きないが、ワープロが普及したことで、複雑さによる使いづらさが軽減され、他の言語では全く不可能な新しい表現媒体へと発展していくように感じている。
このように、私達の日本語は表意文字と音節性が同居するlogosyllabaryと呼ばれる体系だが、syllabogramとlogogramが仮名と漢字に完全に分かれている点がこれまで誕生したlogosyllabaryとは全く違っている。事実、このような複雑なlogosyllabary と呼べるWriting体系は、現在はアルファベットで完全な表音文字を使っている地域でも、歴史の一時期で使われていた。このような例の1つとして、最初は絵文字だけで表現されていたエジプトヒエログリフ表記体系が、が表意文字とそれから生まれたsyllabogramの両方を同時に使用するWritingに発展したことを前回紹介した。ただ注意が必要なのは、このような言語の場合同じ文字がsyllabobramとしてもlogogramとしても使われる点だ。実際、通常logosyllabaryと言うときは、同じ文字を表意と表音節に使う体系を指し、仮名と漢字が明確に分かれる現在の日本語は全く特殊だ。日本語で言えば、万葉仮名が使われた時代のWriting体系がまさにこれに当たる。
このような同じ文字をlogogramとsyllabogramの両方に使うlogosyllabaryでは、必然的に同じ文字に異なる読みが対応するため、混乱を来す。従って、logosyllabary を解消して、syllabaryへと変化しようとする強い力が働く。幸い既に述べたように日本語は表意文字と表音説文字が全く異なる独自のlogosyllabaryを生み出したため、曖昧さの問題が解消され、logosyllabaryが存続できた。個人的には、他のlogosyllabary体系と区別する意味でintegral-aggregram(私の造語で統合的合体体系)と言っても良いのではないかと思っている。

図1楔形文字の進化。 形の変化だけでなく、実際には最初、「頭」を表すsagとして読まれていた物が、頭という意味を持たない音節として使われるようになる。このため、途中段階では、同じ文字が「頭」と1つの音節の両方に使われるようになり、表記上曖昧さが発生することになる。
Pictogramから発して、logosyllabaryへと発展したエジプトのヒエログリフや、メソポタミアの楔形文字によるlogosyllabary体系については既に述べたが、図1はpictogramから始まる楔形文字が、徐々に抽象化されて、シュメールの楔形文字へと発展する様子を示す表でAncientscripts.comと言うサイトより引用している(http://www.ancientscripts.com/sumerian.html)。
図に示された楔形文字は、従って表意文字と、表音節文字の両方に使われていた。すなわち万葉仮名と同じで、最初は表意文字として生まれた文字の一部が、その読みだけを抜き出して音節文字として使われている。この変化を促した最大の理由は、シャンポリオンのヒエログリフについての解読でも述べたように、音で表現される支配階級に関わる名前や場所を表記する必要があったからだと言える。これは万葉仮名でも同じで、残っている万葉仮名の最も古い記録(5世紀)では、人名や地名を書くのに用いられている。
しかし、日本語と異なり、ヒエログリフも、楔形文字もその後表意文字としての使用をやめ、syllabary そして完全な表音文字アルファベットへと変化していく。
なぜ日本語では残ったlogosyllabaryシステムが他の言語ではアルファベットへと変わってしまったのか?一番大きな要因が、logosyllabaryの問題、すなわち同じ音を表す文字がいくつもでき、また1つの文字に違う音が対応する(万葉仮名時代を考えてもらうとわかりやすい)と言う問題が、日本語では漢字と仮名に分けることで解決されていたからと言える。一方、ヒエログリフや楔形文字は、logogramとsyllabogramに対応する表記法を改めて開発する事が難しく、この問題を解決するため残された唯一の方法は、logogramとしての使用を減らしてsyllabogram専用の文字へと変化させる以外なかった。
実際には日本語でもカナが発明された当初は、漢字の使用を減らす傾向が強く見られる。例えば古今和歌集はひらがなだけで表記されているし、枕草子などの文章も今と比べるとずっと平仮名が多い。楔形文字やヒエログリフでは時代が進むと、表意文字の数はどんどん減り、表音説文字だけが残っていく。この結果、記録に残る文字の種類は急速に減少する。
このlogosyllabaryからsyllabaryへの変化をさらに加速させたのが、ヒエログリフや楔形文字が様々な言語で採用され、異なる地域や民族の自由な発想で便利なWritingへの変化が推し進められたことだ。言語と異なり、文字は言語を書き残すためのテクノロジーと言える。従って、どの地域でも便利なテクノロジーとしてそれぞれの言語に合わせて利用した。特に、pictogramやlogogramは話している言語に関わらず使うことが出来る(高校で習った漢文の読み方を考えれば分かる)。その後、それぞれ独自の言語に合わせてlogosyllabaryへと発展させることが可能なことは、仮名の歴史からよくわかる。
実際、最初シュメール人の文字としてスタートした楔形文字も、アッカド語、エラム語、ヒッタイト語など多くの言語に利用され、それぞれの言語で独自に変化していく。漢字が日本語、韓国語、ベトナム語に利用され、独自に発展したのと同じだ。その過程で、writing体系を複雑にする元凶といえる表意文字は加速度的に整理され、日本語の仮名と同じ表音説文字へと変化していく。特に他の言語に利用され始めると、表意文字を整理したあと残る表音説文字は、それぞれの言語で独自の発展を遂げることになる。そのため、楔形文字と言ってもシュメールとは全く異なる、Writingが出来上がる。

図2 アマルナ文書
フリル人の王国ミタンニで使われていた楔形文字。表意文字は消えて、日本語の仮名に対応するsyllabogramだけで出来ている。
Wikipediaより
表音文字が失われて表音節文字のみで書かれた例が、ユーフラテス川上流に形成されたフリル人の王国Mittani語で書かれたアマルナ文書だろう。(図2)。この文書はMittaniの王女がエジプトのアメンホテップ3世と結婚するための条件が書かれており、エジプトのアマルナで出土した。
さらに、楔形文字が記号として拝借されただけで、全く異なるコンテクストで利用されている例も知られている。現在の北シリアの地中海岸に位置し、様々な文化が交わる王国ウガリットで使われていた文字で書かれた文書では確かに楔形文字は使われていても、本来のメソポタミア楔形文字とはほとんど無関係と言っていいWritingが生まれている。文字の種類は合計30しか存在せず、多数の文字を必要とする(例えば日本語の場合(いろは47文字+濁音などで23文字で70文字が必要)syllabaryから新しい表記体系に変化したことがわかる(図3)。

図3 楔形文字を使うウガリット文字 Wikipediaより。
この表から分かるようにウガリット文字は現在用いられているアルファベットとは異なり、子音に対応する文字だけが存在する。実際どう発音されていたのかはよくわかっていない 。
ウガリットは様々な文化の十字路に当たり、ヒエログリフも含め様々な文字が書かれたタブレットが出土している。従って、ウガリット文字は楔形文字で書かれていても、本当はセム語や、フェニキア文字に近いと考えられている。実際、上の表に示した順番を、セム語や22文字からなるヘブライ語と対応させることが出来る(次回)。このように、ウガリットをはじめとする地中海沿岸の国家では、盛んに文化交流をくり返しながら新しいWriting を開発するための様々な実験が行われていたと考えられる。
そしてその中から、エジプトのヒエログリフをルーツとする、フェニキア(セム語)文字が誕生する。ただ、これはまだ子音と母音が分かれた、完全なアルファベットとは言い難く、完全な表音文字の誕生はギリシャ文字誕生まで待つ必要があった。次回はフェニキアのセム文字からギリシャ文字までの過程を見ることにする。
[ 西川 伸一 ]
2018年10月1日
前回、楔形文字による表音節文字(syllabary)が、多くの文化が交わる国際都市ウガリットで30種類のサインに整理されたウガリット文字へと発展したことを紹介した。前回の記事を開いてもう一度ウガリットのアルファベットを見て欲しい。最初の’aに相当する文字を除外すると、全てが子音である事に気付かれるだろう。現在は誰も使っていない、遠い過去のウガリット文字が、本当に子音だけを表現しているのかなぜわかるのかと訝しく思われるはずだ。日本の仮名のようなsyllabogramと考えてはなぜだめなのか? 文字が子音だけで書かれたとすると、正確に読むことは出来るのか? など疑問は尽きない。
実際には、ウガリット文字には子音に対応するサインしかなかったことを疑う人はいない。というのも、この文字に多くの親戚があり、それらは同じように子音を示すサインだけを使っている。また現在でも子音だけから出来た文字を使っている言語がある。その一つの例が、ヘブライ語だ。図1にヘブライ語のアルファベットを示すが、最初のalep(アポストロフィで表されている)は別にすると、全て子音だけだ。

図1:ヘブライ語のアルファベットと読み。(Wikipediaより)
子音だけからなる表記体系があるのは分かったが、実際にはヘブライ語ができない私には、本当に子音だけで文章を正確に理解する事が出来るのかは判断できない。しかし少し考えてみると、この方法に慣れさえすれば、文章全体が表現しているコンテクストから子音の次に来る母音はおおむね判断ができ、間違いなく読むことが出来るように思える。例えば日本語で「こんにちは:KONNICHIWA」から母音を省いてKNNCHWと書いみよう。最初はわからなくても、慣れてさえくれば読めるのではないだろうか。
もちろん母音が指示されないと、当然間違う確率も高まるはずだ。この問題を避けるため、現在のヘブライ語では正確を期すため、母音に対応する記号を文字の下に小さく書き足して曖昧さを避けている(図2)。

図2:ヘブライ語の表記に母音が足されている例。
文字の隅に小さく母音の記号が足されることで、正確な読みが可能になる。(Wikipediaより
このように子音だけからなるWritingが紀元前から現在まで存在してきたことは理解していただけたと思う。このような表記体系は、子音アルファベット(consonant alphabet)と呼ばれており、現在使われている言語では、ヘブライ語やアラビア語がその代表だが、これらは全て前回説明したウガリット文字と兄妹の関係にあるといっていい。
図3はこれらの文字の系統関係を示した図で、わが国を含む東アジアの文字を除くと、現在世界で使われている文字のほぼ全てがこの系統の中に含まれる。
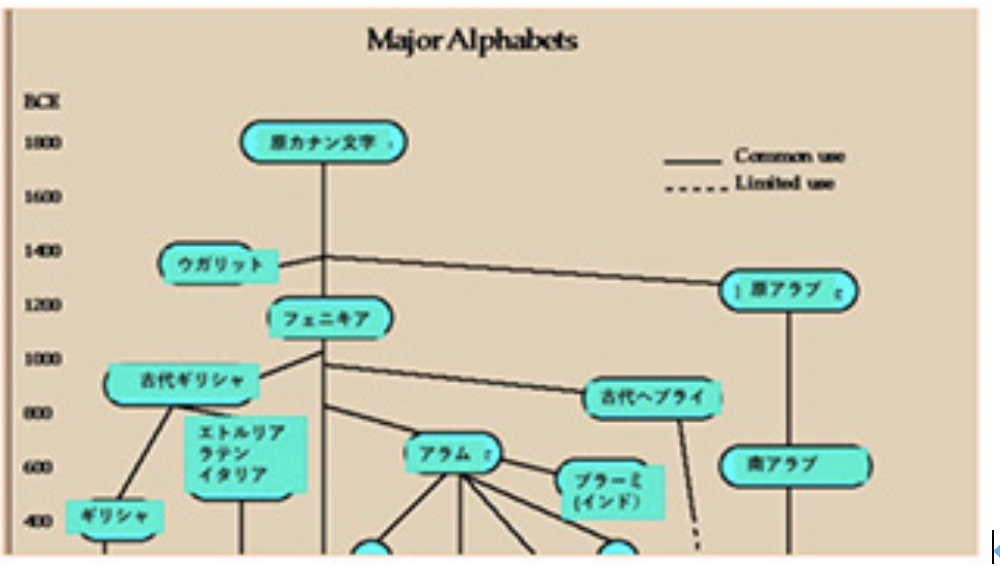
図3 ヒエログリフを起源とする原カナン語から派生した様々な言語。現在使われているほとんどの凶器体系が含まれる。(Ancientscriptの図を改変)
この図が示すのは、現在世界中で使われている表記体系は原カナン文字に由来する点だ。この原カナン文字は、カナンで発見された文字で図4に示すようにエジプトのヒエログリフから、原シナイ文字を経て発展してきた文字で、ヒエログリフと現在のアルファベットをつなぐ役割をもっていたといえる。
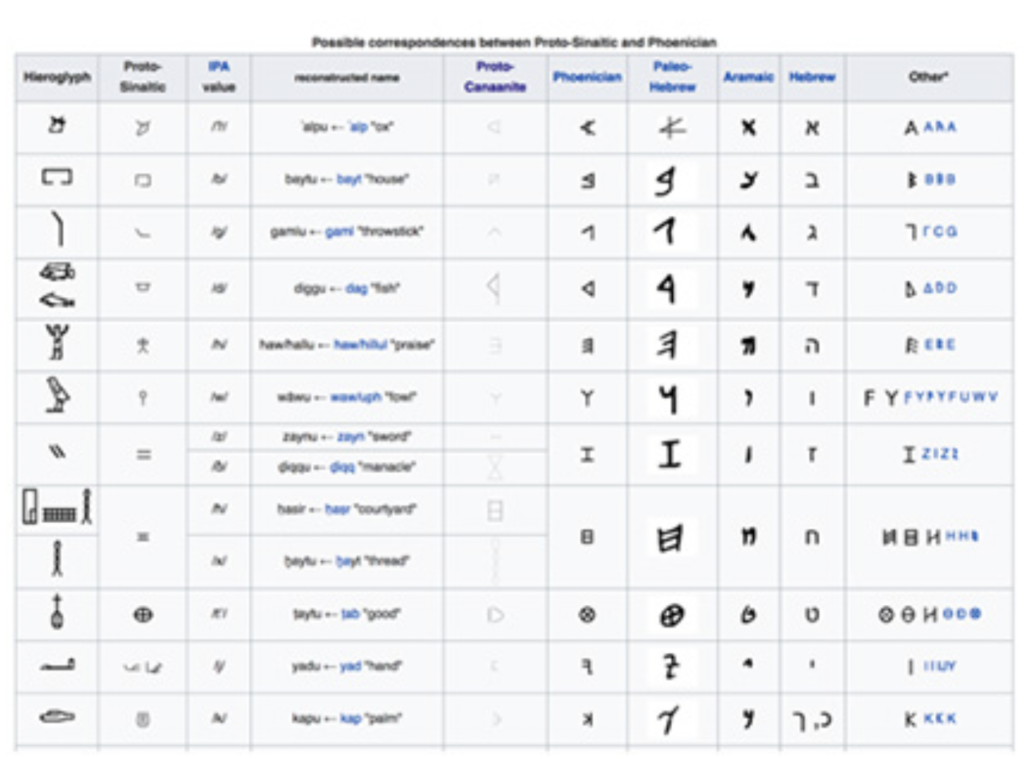
図4 ヒエログリフに由来する様々な文字。(Wikipediaより)
図4に示すように、恐らく多くの言語でおなじ様な表記体系が使われていたと思われる。実際、ウガリット文字は楔形文字を使っているが、おそらく粘土板に書く必要があったため楔形文字の特徴を利用しただけで、お手本は原カナン文字や、原シナイ文字にあったと考えて良い。このため、ウガリット文字も含めそれぞれの文字をアルファベットとして並列に並べることが出来る。すなわち、どのようなサインを使うのかは、記録する媒体に応じて自由に決められたと考えられる。
それぞれ話す言語は大きく違っていたはずなのに、なぜ母音を省く表記体系がこれほど普及したのかは面白い問題だ。これは私の勝手な想像だが、母音の数は多くないのに、記載するとなると子音と同じ回数だけ文章内に現れる。石に彫るにせよ、粘土板に刻むにせよ、あるいはパピルスに書くにせよ、母音をそのたびに書き残すのは手間もかかるしお金もかかる。このため、極力記録のための文字数を減らしたいという要求があったのだと思う。こんな単純な理由で、後で理解できればいいと子音だけを使ったアルファベットが普及したのではないだろうか。
漢字が朝鮮、日本、ベトナムでそれぞれ利用されたように、便利な発明は文化の交流と共に、急速に普及した。事実紀元前1000年前後にはエジプトから現在のシリア、イランまで、盛んに交易が行われ、人的交流があった。前回述べたように、ウガリットでは、様々な文字が書かれた粘土板が出土していることは、ウガリットに文字を含む様々な文化が入ってきていたことを示している。やはり前回紹介したアマルナ文書からわかるように現在の北シリアにあったミタンニ大国の王女と、エジプト王朝の王子が結婚していたと言うことは、紀元前1400年頃には王国間の交流が盛んに行われていたことを示している。
このような様々な民族間の活発な政治的、経済的、文化的交流の中で、レバノンから北シリアを中心に地中海全域で活躍していたのがフェニキア人で、交易を通して様々な文化を導入する過程で、交易に適したわかりやすい線文字、フェニキア文字を開発した。ただ、図4に示すように、当時セム語を話す民族が用いていた文字は全てよく似ていたと言える。その中で、フェニキア文字がアルファベットの起源とされている理由は、フェニキア人とギリシャとの関係がもっとも深かったためだ。その結果ギリシャ人はフェキキア文字を手本にして、全てのアルファベットの原点ギリシャ文字が考案される。
実際フェニキアという名前の由来だが、フェニキアとは土地の名前ではなく、ギリシャ語で「赤い手を持つ人」と言う意味を持っている。すなわちギリシャ語で呼ばれるほど、ギリシャとの関係は深かった。なぜ「フェニキア:赤い手」と呼ばれたのかというと、レバノンから北シリア沿岸にかけて生息している貝から得られる貴重な染料を重要な商品として扱っていたため、手が何時もその染料でそまっていたからだとされている。
このように、ギリシャとの交流が盛んだったフェニキアを介してギリシャに原カナン文字由来の表記体系がもたらされる。そして、この子音アルファベット文字に母音を加えた全く新しいアルファベットが考案されることになる。
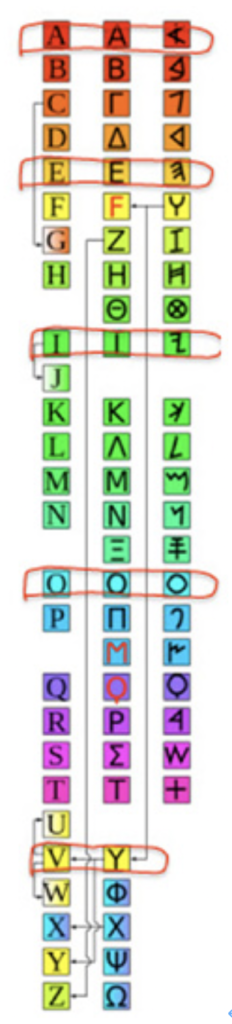
図5 英語(左)、ギリシャ語(中央)、フェニキア語(右)のアルファベットを比べた図。囲った文字が、ギリシャ語や英語で母音として使われている。
図5にフェニキア文字(右)とギリシャ文字(中央)、そしてアルファベット(右)を並べて比べてみたが、英語のアルファベットもギリシャ文字のアルファベットも6文字、増えているのがわかる。線で囲った文字が、ギリシャ語や英語での母音になるが、フェニキア文字の発音の中から、母音に近い物を取り出し使っている点に注目してほしい。すなわち、alephをA、heをE、yodhをI, ayinをO、そしてフェニキア文字では最初の方に存在していたY=wawをUの母音に対応する文字として使うようになった。すなわち、現在のヘブライ語で意味を取り違えないために使われている新しい母音の記号を足す方法ではなく、フェニキア文字が本来持つ音を利用して(rebusに近い)、母音を持った表記体系を新たに作り直していることがわかる。
ギリシャの表記体系は、フェニキア語のよくわかる一人のギリシャ人によって、約紀元前800年頃に考案されたと考えられている。ここで気になるのは、それまで子音だけを表記する体系で済ませてきたのに、ギリシャに渡った後、どうして母音を持つアルファベットへと再構成されたのかという点だ。
勿論ギリシャでは、時間も労力もあり、わざわざ母音を省く理由がなかったのかもしれない。あるいは、ギリシャ語では母音を省くことの不都合が、他の言語に比べると多かったのかもしれない。
他にも様々な理由を思いつくことが出来るが、一番魅力的なのがWritingの著者Barry Powellが唱える説だと思うので、最後に紹介しよう。
ギリシャ文字が出来たと考えられる紀元前8世紀は、ギリシャの叙事詩を生んだホメロスの生きた世紀だ(図6)。オデッセイアやイーリアスを読んでいなくとも、アテナやアキレス、ポセイドンを知らない人はいない。すなわち、ギリシャのみならず、ヨーロッパ文化の原点にある一大叙事詩だ。

図6 ホメロスの胸像とオデッセイア(Wikipediaより)
驚くことに、このホメロスの壮大な叙事詩も、最初は口ずさまれていたらしい。すなわち、彼の叙事詩は記録されることを想定せず作られた口語体の文学だったと言える。実際、あらゆる文化でこのような口述の物語が存在する。我が国の語り部、ヨーロッパの吟遊詩人などはその例で、おそらく大部の内容をしっかり頭に入れるために、メロディーを持つ詩として構成されたのだろう。しかし、今残っているこのような文学は、幸いにも文字に記録されることで消滅を免れた。
当然ギリシャでも重要な叙事詩を記録しようということになったが、そのためには口語を正確に写す必要がある。これは、これまで行われてきた伝えたい内容だけを記録するのではなく、話されていることをそのまま写すことに他ならない。Powellはこの語られた叙事詩や詩をそのまま写すために、母音を文字として表すギリシャ語が誕生したと提案しているが、少なくとも私にとっては最も魅力的な説だ。
この大変革の結果、ギリシャ語の表記体系は、少しづつは変化しても、現在まで生き残ることになった。またそれだけでなく、ローマで現在のアルファベット表記体系へと作り変えられ、現在多くの言語で用いられるアルファベットのルーツとなった。
次回は、文字の機能について考えて見る。
[ 西川 伸一 ]
2018年10月15日
さまざまな文字の歴史をたどってみると2つのことがわかる。まず、Writing systemの進化が、話しことばに使われる音素をそのまま表現できる技術へと進んできたこと。そして、現在でも6000種類存在し、過去にはさらに膨大な数が存在した言語自体と違って、文字の起源はほんの一握りのルーツへと遡れることだ。
現在使われているWriting systemをたどると(図1)、世界で使われているアルファベットや子音アルファベットは全てエジプトのヒエログリフ由来の原カナン文字へと遡れる。実際、漢字を最初writingに用いた中国語、日本語、韓国語だけは現在も独自の文字を使っているが、他の言語はローマ字のアルファベットも含めたフェニキア文字由来の子孫に置き換わっているといっていい。ヘブライ語、アラビア語やペルシャ語は一見アルファベットとかけ離れた文字で、表子音文字だが、既に説明したように全て原カナン文字に由来する。
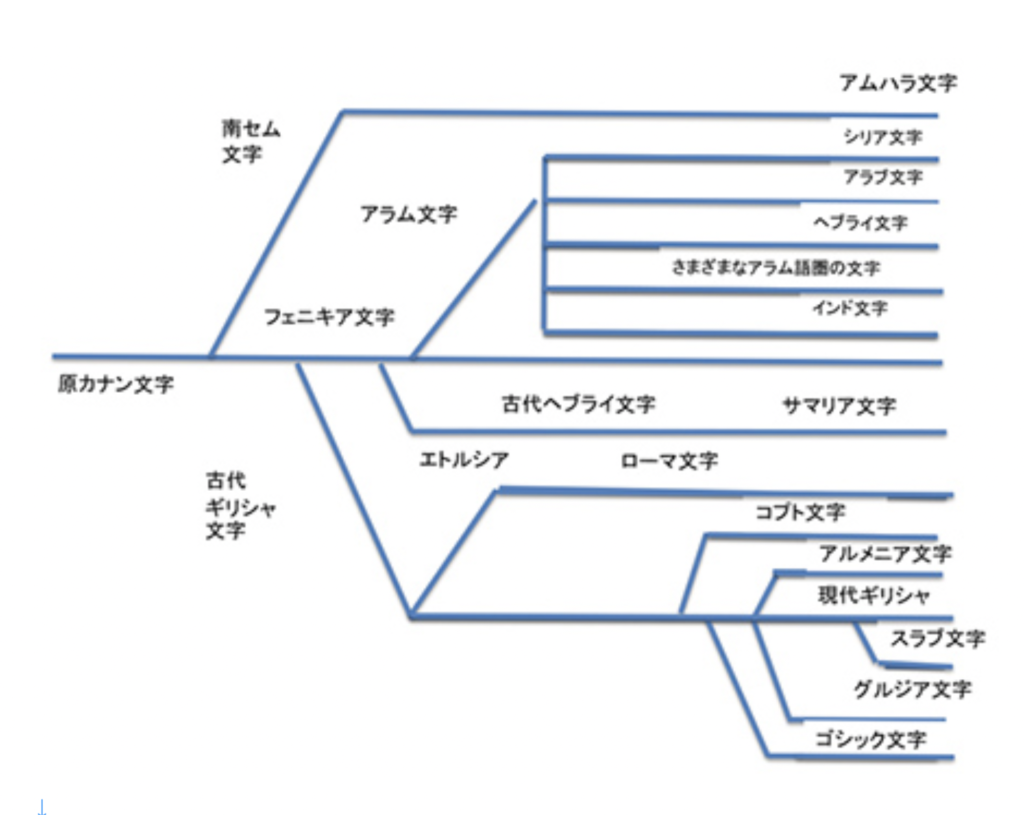
図1: 現在使われている文字の系統樹 Blatner博士の講義(https://www.blatner.com/adam/scriptology/3-Mid-History/3-midhistory.html)を参考に作成。
同じ様に、インドから東南アジアにかけては、極めて多様な、一見完全に独立したかに見えるサインを用いているが(図2)、これらは全てブラーフミー文字に由来しており、ほかの文字と共通祖先を持っている。

図2 インドから東南アジアにかけて独自に進化した、ブラーフミー文字由来の文字(Wikipediaより)
図1からは除外されているが、漢字だけが独立して生まれたとは考えにくいと、シュメールの記号にその起源を求める研究者もいる、もしそうだとすると結局世界の全ての表記法がつながっていることになる。これまで漢字の起源は甲骨文字という常識を覆し続けている新石器時代 由来のサインを見てみると(図3)、個人的には確かにシュメールに近いかもしれないという印象を持つ。もちろん漢字は全くオリジナルな表記法だとしても、当時既に世界レベルの交流があったと考えると、文字という便利な道具があることを知り参考にした可能性は大いにある。

図3 半坡陶符は土器に残されている、新石器時代の文字に近い記号と考えられている。これ以外にも、龍山文化遺跡から甲骨文字とは違う記号が出土している。(Wikipediaより)
このように、各Writing systemが少数の起源(ひょっとしたら一つの起源)に絞れるのは、言語とは無関係に、文字を道具としてそれぞれの言語に導入できるからだ。しかし、起源が共通と言っても、それぞれの文字は独自に進化しており、どんなにアルファベットが合理的な表記法だと言っても、それに収束する物ではない。現在実際に使われているWriting systemは、アルファベットを用いる完全な表音文字、アラビア文字、ヘブライ文字のように表子音音節文字、そして音節文字や表意文字、日本語のようなその混合まで、いまだに進化の過渡期をみる有様だ。実際、個人的には過渡期だと思っている。というのも、ほとんどの人が文字を使うようになる現代世界では、間違いなくアルファベットの様な完全な表音文字へ収束させようとする圧力は強くなると思っている。例えば我が国や中国では、文字の使用が大衆化する過程でアルファベットの導入が真剣に考えられたことがある。また、それぞれの言語にとってより便利な表記法の開発はそう難しいことではない。古くは日本語の仮名がそうだし、新しくは朝鮮語のハングルがその例だ。その意味で、まだ完全な表音文字を用いていないWriting systemは、アルファベットへと収束するのではと予測している。さらにグローバル化に伴い、アルファベットがもう一つの表記法として、各言語の公的表示と共に利用されている(図4)。今後どこまでアルファベットが進出するのだろうか?

図4:イスラエル、中国、サウジアラビアの道路標識。(Wikipediaより)
ただ、アルファベットの勢いをとどめる大きな要因が、Writing systemが本来持っている社会の階層性を維持し、差別を助長するという特徴だ。Writing systemの究極の目的が話し言葉の記録だとしても、大衆化を目指して文字の歴史が進んだのはほんの最近のことで、長い間限られた階級の、限られた目的のために、Writing systemは発展してきた。これが現在のwriting systemの多様性の背景にある。言い換えると、それぞれの文字はその使われ方に依存して発展していると言える。もう一度Writing systemの歴史をおさらいしてみると、文字は社会の階層化と共に発展し、また階層化を拡大する、差別の原動力として働いたことがわかる。
まず、トークンが粘土板の記号として使われていく過程は、農耕による社会の階層化と一致している。すなわち、トークンやそれに由来する記号は所有する財産の覚書として使われて来た。この時代は絵文字だけで表記が行われていたと思われるが、狩猟採取民の平等社会が完全に崩壊したことをうかがわせる。
次に、王や神官などの権力者の様々な覚書として文字が用いられるようになる。おそらくこの時、意味を表す文字がrebusの原理で音を表す文字としても使われるようになる。というのも、例えば正確に人名や地名を記載するためには、音を表現することが必須になる。その結果、漢字を導入した時代の日本語のように、表意文字とその一部を表音文字としても使うというlogosyllabaryとよばれるWriting systemへと発展する。同じ記号が意味と音の両方の表現に用いられるため、当時のwriting systemはきわめて複雑で、そのため文字を扱うテクノクラートを養成し、書記としてかかえることが必要だった。実際、エジプトやメソポタミアの遺跡からは、数多くの書記の像や絵が発見されている(図5)。

図5 エジプトの書記像。(Wikipediaより)
このように、一旦Writing systemが王や神官を頂点とする階層性を維持する装置として機能し始めると、大衆化はほとんど望めないことがわかる。
そんな中でこのような階層性を乗り越えて、Writing systemの一定の大衆化を促した1つの要因が、地中海を股にかけて行われた交易で、この結果その後のWriting systemの手本となるフェニキア文字がこうして完成したのも頷ける。
Writing
systemの大衆化を促した2つめの要因は、神の下での平等を唱える宗教、ユダヤ教の誕生だろう。旧約聖書には有名な出エジプトの物語が記載されているが、考古学的にはこのような事件が起こった痕跡はなく、作り話として考える学者が多い。しかし、このような統一の物語が必要だったのは、最初のイスラエル王国が紀元前1000年頃に成立するまで、ユダヤ人は分断され、常に民族としての統一を求めていたからだろう。さらに、紀元前587年にはユダヤ人はバビロンに捕らわれることになる。すなわち、階層性が完全に破壊されたなかで、民族の宗教が書かれた教典として示されることで、あたらしい大衆化したWriting systemの機能が生まれる。
これはキリスト教も同じで、支配者を中心とした階層社会ではなく、神の前に平等な信者にWriting system が必要とされる。この時使われたのがすでに大衆化が進んでいたギリシャ語で書かれていたことは納得できる。
すでに書いたように、writing systemが階層性の維持としての役割を捨て、大衆化したもう一つの要因が、文化の共有だった。すなわち、口述されていたギリシャの叙事詩を写し取る目的でギリシャ文字が考案され、その後のアルファベットの原型になる。既にギリシャはポリスを中心とする民主社会を形成しており、構成メンバーが平等な市民だったことを考えると、市民の誰もが学習しやすい正確な音素の表記法がギリシャで完成したのも頷ける。
日本語でも仮名、特に平仮名は、ギリシャ語と同じで音で詠まれる文学を写すという機能を持っていた。元来和歌はギリシャの叙事詩のように詠まれる文学だった。従って、漢字のような表意文字のみでは表現できず、音を移すことのできるWriting systemの開発が必要だった。このため最初の万葉集では万葉仮名が使われるようになる。そして、おそらく文学に使われる日本語の読みを表記するため、平仮名が考案されたと考えられる。勿論最初から和歌の表記に使われたわけではないが、公的な文書に平仮名が最初に使われるのは、言うまでもなく勅撰和歌集、古今和歌集で、このことからも詠まれる文学の表記が重要な平仮名の機能だったと言える。
さらに平仮名がギリシャ文字と似ているのは、階層性を壊す力を持っていたことだ。ギリシャの市民社会と比べると、当時の日本に市民社会が成立していたとは言い難い。平仮名が誕生してWriting systemが大衆化したとは到底言えないが、しかし男女の差別は完全に平仮名の誕生で解消されたように思える。古今和歌集には小野小町が六歌仙として選ばれており、他にも伊勢など女流歌人も差別されることなく選ばれている。これを大衆化と位置づけても良いのではないだろうか。実際、平仮名により文字の大衆化がすすみ、江戸時代にはなんと識字率が40%近いという驚異の数字が達成される。

図6 古今和歌集(Wikipediaより)
一方各民族の政治や文化の階層性や差別性が、例えばアルファベットのような便利な表記法へとWriting systemが収束するのを阻害することも多い。イスラム圏でもトルコやインドネシアでは、表子音文字であるアラビア文字は文字の大衆化を阻害するとして、早くからアルファベットに転換している。しかし、他の国ではこれまで通りアラビア文字が民族や国家のアイデンティティーを守るため使い続けられている。このように、恐らく言語の重要な機能は、民族や文化のアイデンティティーを守ることだ。勿論この機能に、言語の大衆化は必要ない。逆に、言語の大衆化が問題になるときは常に、文化的アイデンティティーは侵される場合が多い。このことは図2に示したブラーフミー由来の表記法の多様性にも反映されている。インドが国家として統一された後、何度もこれらの表記法を統一しようと試みられたが、現在もなおうまく行っていない。これもWriting systemと言語を一体化させることが文化や民族のアイデンティティーに必須であるとする考えに基づいている。
このようにWriting systemが様々な機能と共に、文化のアイデンティティーを守る機能を持っていることを考えると、日本人として自分で使いながらもほとんど理解できなかった「なぜ3000年以上も変わることなく、漢字のみを使うWriting systemが存続したのか」という答えが見えてくるように思う。私は中国語が全く出来ないが、漢字とその読みの成立を考えると驚きを隠せない。Barry Powellの「Writing」に記載されていた例で考えてみると、日本語では「安」と「案」をともに「アン」と読む。まず安だが、家をあらわす「うかんむり」の下に女がいると、平安があるという意味で成立した漢字だ。そこに、「木」を足す事で中国では机と同じ意味になる。中国語では2つの文字を実際にはアクセントを違えて表現するが、それで本当に意味が通じるのか不思議だ。実際日本語では、1つの文字をそのまま読むことはなく、安心、案件といった熟語にするか、案ずる、安んじると助詞をそえて意味を正確に伝える。中国語も時代と共に外来語も含め話し言葉はますます複雑化している。しかしこのような原理のWriting systemでは、どんなに漢字の数を増やしても、正確なコミュニケーションが可能なのか、中国語を知らない私でも心配になる。

図7 矛盾という単語のいわれを説明した漢文。カナが振ってあり、完全に日本語として理解できる。(Wikipediaより)
しかし漢字の素晴らしさもある。例えば、Wikimedia commonsから引用した「矛盾」というタイトルの漢文は(図7)、完全に日本語化して読むことが出来る。もちろん、中国語や朝鮮語としても、実際には英語であってもしっかりと読むことができるだろう。すなわち話し言葉と対応させる必要のない国際的表記法が完成している。全くの私見だが、このことから漢字は歴史的に大衆化への意図を全く抱くことなく発展したWriting systemと言えるかもしれない。言い換えると、中国語の役割はヨーロッパの学術や宗教の共通語ラテン語と同じ役割を担ってきたと考えられる。すなわち、話し言葉との連続性を犠牲にしても、中華の伝統を守り、伝え、なおかつ国際的なWriting systemとして維持されてきた。このおかげで、我が国や朝鮮、また周辺の様々な国で、仏教、学術、文学などを共有することが可能になった。
だからこそ、20世紀に入って中国がWriting の大衆化の必要に駆られたとき、アルファベット表記のピジンが開発され、また外来語を訳し直さずそのままrebusの原理で表記し、漢字を思い切って簡易化したWriting systemが瞬く間に形成される必要があった。そして「中国語の共通文字体系である漢字の歴史は古い。」という日本語の文章は、本来の中国語に近い台湾で用いられる中国語(繁)では「作為中文的共通文字體系的漢字的歷史舊」表記されるのに、簡易化された中国語(簡)では「汉字的历史,是一个普通的中文字母系统,是古老的」と、かなり話し言葉に近い形で表記されるようになっている。
このように、社会的、文化的にWriting systemが果たす機能を考えると、差別化、階層化を維持する役割と、新しい大衆的情報としての役割の間で、相反をくり返しながら発展してきたと思う。この意味でWriting systemは言語そのものより人類の文明に大きく寄与してきたように思う。次回は、文明の原動力としてのWriting systemについて考えたい。
[ 西川 伸一 ]
2018年11月1日
前回、Writingが、基本的には道具として多くの人が使いやすいように進化し、最終的に最も単純化され、最もフレキシブルだった表音文字アルファベットがほとんどの言語を席巻したこと、しかし、言語自体が持つ文化を担うという役割のため、writingの多様性が維持され現在にいたることをざっと見て来た。
これでいよいよ情報としての文字についての議論は終えるが、最後に文字の機能をたどりながら、Writingと文明や文化の関係を復習するとともに、すこし実証性を犠牲にしても、個人的な考えを積極的に交えて、Writingが文明や文化の発展にどう関わってきたのかを2-3回にわけて考えてみたい。
1、記憶とWriting
音節を用いる言語の誕生から文字の誕生までを考えてみると、ともに私達の記憶の限界を拡大することを1つの目的として生まれてきたことがわかる。既に述べたように、視覚を通して私達にインプットされる情報量は膨大で、しかも記憶には多くの脳領域がかかわる。従って、記憶を呼び起こすとき、脳内に形成される別の表象と連合させることで、記憶を高めることが出来る。この脳内の連合対象として、記憶しやすい音節の表象を用いるようになったのが、私達の話しことばのルーツになる。おそらく、最初は音を個人的メモとして記憶に使っていたと思われるが、この音節に対応する表象を共有することが出来ると、コミュニケーションのための音節を用いる言語が誕生する。
Writingの歴史を見ていて面白いのは、同じ様にWritingのルーツが、個人の覚え書きとして使われたトークンにあるとされている点だ。図1にルーブル美術館に展示されているトークンとそれを納めていたBullaeの写真を示すが、これらは自分の財産を管理するために使われたと考えられている。前に並べて展示されているのがトークンで、同じ形のトークンは同じ所有物(例えば麦一袋)に相当する。大型のトークンは、例えば5袋を表すのかもしれない。倉庫に行って数えれば済む話しだが、盗まれて数が減る危険があったのだろう、すなわち、一種の記帳の目的でトークンがつかわれている。例えば「記録しているから盗んでもわかるぞ」と言ったコミュニケーションの意味はあったかもしれないが、原則は自分のための覚え書きと言える。ただ、音節言語により増強した記憶とくらべた時、トークンが可能にする記憶は正確さ、持続時間の点で、質的大転換が起こったといえる。

図1ルーブル美術館に展示されているトークン
このようにトークンは記憶のための道具で、誰かがトークンを発明してその便利さがわかると、他の人たちも使い始め、広がったのだろう。文字の誕生後もトークンは使い続けられたようだが、最初個人の手作りだったトークンも、専門の職人により造られるようになる。恐らくその時、それぞれのトークンが表す意味も他の人たちと共有されることになり、交易にも使われたりすると、一種のコミュニケーションの道具に変化する。すなわち、Writingの誕生にグッと近づく。
トークンを文字のルーツとして見てみると、Writingに備わった、言語とは違う2つの重要な性質が見えてくる。その1つが、トークンや文字によって、記憶を正確に、長い期間保持する優れた道具ができた点だ。そしてもう一つが、このような記憶道具は、社会の階層化を反映していることだ。おそらく、私有財産、交換、分業といった文明の下部構造がこの時生まれ、私有財産が文字への道を開いたと言っていいだろう。
事実、トークンは約1万年前に発明されたと考えられるが、だとすると音節を用いた言語誕生から何万年も、このような正確・長期の記憶道具の必要性はほとんどなかったことになる。実際、現在でも未開の狩猟採取民ではほとんど表記法が発達していない。その日暮らしで財産が残らない狩猟採取民では、優秀なリーダーとそれに従う仲間といった関係はあるにしても、文明のための下部構造が形成されない。
トークンは農耕や牧畜にすこし遅れて誕生したと考えられる。農耕も牧畜も、定住と同時に、私有財産、分業、交換を生み出したはずだ。そしてトークンが示すように、わざわざ道具まで作って家の中に置いておかないと管理できない量の財産が発生した。また、トークンはコミュニケーションの道具としても役立ったのかも知れない。すなわち、持てるものが自分はちゃんと管理しているから、盗んでもすぐわかるぞと示す意思表示だ。このように、トークンは記録を必要とする貧富の差が生まれる社会構造が存在したことの証拠になる。
その後トークンを粘土板の表面に押し付けて記録する、文字に近づいた表記法が考案されるが、これらもやはり個人の覚え書きの域をまだこえていない。すなわち、社会で共有される表記法とはならなかった。というのも、これらの覚え書きには、伝えるというコミュニケーションの側面が欠けていた。これが生まれるためには、さらに大きな社会と強大な権力を中心とした階層性が必要だった。
2、Writingによる最初のコミュニケーション:権力の誇示
トークンを押しつけた粘土板を手始めに(図2)、絵文字を並べたWritingが発達したと思われるが、おそらく絵文字のみで表記が行われていた時代でも、Writingのコミュニケーションとしての役割は限定されて、なかなか覚え書きの域から出ることはなかったのではないだろうか。 実際、このような段階はProto-Writingと称されており、ここに書かれている文字は多くの人と情報を共有するために使われていたのかどうかは怪しい。例えば、財産管理なら何が書いてあるかを知られない方が良い場合すらある。

図2 メソポタミアの粘土板
財産管理の記録。ルーブル博物館で撮影。
一方、言語の最も重要な機能は、その誕生から現在までコミュニケーションだ。従って、言語と独立に絵文字や象形文字が発達したとしても、なかなか覚え書き以上に発展することは出来ないと思う。したがって、Writingが正確で永続的に情報を伝えるコミュニケーションの道具として発展する為には、話し言葉、即ち音をそのまま表現できる文字の誕生が必要だった。即ち、音節による言語の限界、すなわち正確さと持続性の限界をWritingにより克服して、永続的に情報を伝えたいと考えた人たちの中に、Rebusの原理(http://www.brh.co.jp/communication/shinka/2018/post_000010.html)、すなわち象形文字や表意文字が持っている音をそのまま借用して利用する言語の表記方法に思い至る人たちが生まれる事になる。これが達成できれば、音声言語の音を写す機能を獲得したWritingには自動的にコミュニケーション機能が付け加わる。
しかしだからといってWritingが、今私達が使っているような意味で一般のコミュニケーションの道具になったわけではない。トークンと同じで、Writingを利用できたのは権力とそれにつながる一部の人だけだった。というのも、最初象形文字が表音Rebusと混在した表記法をマスターするのは子供の時からの教育が必要だった(我が国ではこれは今も同じだが)。これには大変な努力が必要で、王と言えども必ずしも読んだり書いたり出来なかったことがわかっている。従って、エジプトでもメソポタミアでも、多くの教育を受けたテクノクラート、書記が必要だった。

図3 エジプトの書記像
ルーブル博物館で撮影。
このように、誰が何を伝えるかという点で、言語とWritingは大きく違ってしまう。まず、話し言葉は、発話者とその近くにいる音の届く範囲の聴き手とのコミュニケーションに使われる。基本的には、気心の知れた仲間同士のコミュニケーション、すなわち対話を中心に使われた。ところが、Writingになると、伝える対象の範囲はほぼ無限に広げることができる。現代なら、Writingは恋人同士の手紙から、不特定多数を対象にする本や新聞の記事、そして法律まで、極めて多様だが、これはあくまでも現在の話で、当時は階層が高い階級が低い階級に伝える、一方的な伝達手段として使われるようになる。
事実、現在残っている最も古い文字は、墓やお棺に書かれた埋葬された人の名前やその人の伝記の記録で、この名前の音をシャンポリオンもヒエログリフの解読に用いた。すなわち、名前を呼ぶときの音を記録したいと言う欲求が、話し言葉を写すWritingの発明を促し、この結果、権力者の歴史(伝記)を、オーソライズされた一つの話として誰にでも同じように伝えることが行われるようになった。その後Writingは、同じように変えてはいけない内容、すなわち行政記録やハンムラビ法典(図4)のような規範の記述に使われるようになる。
異論もあるかもしれないが、話し言葉を写すWritingは、権力者の言葉を権威化、正当化する為に発明されたと言っても過言ではないと思う。言い換えると、Writingはその使用を許されたものだけに、さまざまな歴史や法(権力者の)を固定的にオーソライズすることを可能にし、それを持たない者との格差を拡大するのに大きく役立ったと言える。

図4 ハンムラビ法典
裏に楔形文字(アッカド語)当時の法律が書かれている。図に示すように、この法が神から授けられた者である事を示す彫刻が書かれている。
ルーブル博物館で撮影。
モーゼがシナイ山で神から十戒をさずかった話も、これによく似ている。
基本的には、伝える内容と、伝える人間を神がオーソライズしているわけだが、実際にはWritingが権力をオーソライズしている。
そして、Writingを使える人と、使えない人の非対称性が、文明の新たな段階、即ち広い国土を持つ文明の誕生と一致している。
エジプト、メソポタミアの時代からWritingをマスターする為には時間がかかった。この為、最初Writingに付与されてしまった階層性は、近代に入って教育の普及により徐々に解消され、Writingを通した会話と言える手紙が一般にも普及する。それでも、例えば作家と読者と言った階層性は、どうしてもWritingから完全に切り離すことができないのは、おそらくこの誕生の歴史を引きずっているのではないだろうか。
最初に現れた文明、エジプト、メソポタミア、中国、インダスなどは、単純に農業社会が拡大してできたわけではない。広い国土を収め(たとえば灌漑)、多くの人口を養えるための、為政者を中心とする階層社会を基盤とする政治体制が必要だった。そして、それぞれの文明で独自のWritingが形成されたことは、Writingが初期の文明を支える重要な機能を帯びていたこと示しているように思える。とすると、Writingが最初から階層性を持っていたことは納得できる。
3、文字が書かれた媒体と階層性
今回はここまでにして、次回は、文字が完全な表音文字へと発展する過程を文明と社会の観点から議論してみたいと考えている。ただ、Writingがより広い階層に普及し、権力の意思を伝える以上の発展を遂げる為には、Writingを支えるテクノロジーの進歩も重要だった点に注目しておきたい。たとえば、粘土板の上に文字を書いた場合、粘土板の重さからみて、気軽に手紙を書くのに使うというわけには行かなかっただろう。この点でも、Writingが最初権力者とそれに連なる人にだけに独占される理由はあった。この状況が、新しいテクノロジーでどう変化し、writingが普及したかについては次回以降の議論になる。
その代わりに、今回は最後に、これまでになかったWritingを支えるテクノロジーのおかげで、今Writingが大きな革命期にあることを述べて終わりたいと考えている。
この革命とは、スマートフォンを用いたメールやソーシャルネットワークの普及に後押しされた変革だ。これまで述べてきたように、話し言葉の基本は、一対一に限るわけではないが会話にある。一方、Writingは手紙という形式は誕生したとしても、会話に用いられることはなかった。ところがどうだろう。スマートフォンが普及してからは世界のどこにいっても図5に見られるように、スマートフォンを使っている人たちを見かけない場所はない。

図5 スマートフォンを使うアラブの女性。(https://pxhere.com/en/photo/1447611より転載)
実際2017年ではスマートフォンの利用台数は40億台を超えたと言われている。おそらく世界の成人の半数がスマートフォンを使用しているといっていいだろう。そしてこのスマートフォンの用途の最大のものは、図6で示したメッセージアプリの画面のように、会話なのだ。

図6 メッセージアプリでのチャット。
(https://www.deviantart.com/wonderland902/art/Text-conversation-with-Big-Brother-454497493から転載)
すなわち、Writingが誕生して4000年、今Writingが言語と同じように会話に使えるようになった。そしてこの中で、EMOJIのような新しい表記法が開発され使われ始めている。このwritingが持っていた階層性を破壊する新しいトレンドが、私たちにどのような新文明をもたらすのか、ぜひ見て見たいものだ。
このようにWritingを支えるテクノロジーはその性質を大きく変化させる。次回は、Writingの普及を促した文明要因について考えて見たい。
[ 西川 伸一 ]
2018年11月15日
前回、文字には最初から、伝える人と、伝えられる人の間にある大きな階層性が内在していることについて議論した。ただこのような階層性を内在しつつも、Writingが文明を変革していく力のある大発明であることはおそらく使った誰にもわかったはずだ。このため、文明や文化の大きな転換期には、常にWritingとそれを支えるテクノロジーが変化してきた。言い換えると、文明とWritingは相互作用を通して互いに変化を促進してきた。
Writingは何に使われたか?
Writingは最初一方的な伝達に使われていたが、その後時代とともに伝える・伝えられるの関係が双方向性を帯びていく。これは文字により書かれた内容を追いかければ自ずとわかる。どの言語でもいいのだが、1994年に阿辻哲次さんにより書かれた「漢字の文化史」を参考にして、漢字を使って何が書かれてきたかを追いかけてみよう。
図1 1994年に出版された漢字の文化史 漢字の歴史についてはこの本を参考にした。
阿辻さんの本によると、亀甲文字が占い目的に使われたことからわかる様に、漢字は最初神への呼びかけとして使われた。その関係で、占い以外に使われる場合も、先祖の霊を弔う言葉を子孫に伝える目的で伝えられていたようだ。しかし、周から春秋時代へと移る頃から(約紀元前1000年)、主君から家臣、親から子孫への訓告といった人間臭い内容へと変化し、さらに国家間の同盟に関する誓約書、荷物輸送時の免税証明書などのような、極めて実用的な文章に使われるようになっている。これと並行して、それまで亀甲や青銅器に書かれていた文字は、石、竹、木片、そして絹などの上に記録される様になる。
主君や家長の訓示は一方的伝達だが、同盟書や免税証明書は、両者の合意を反映させる必要があり、決して一方的に書かれるものではない。また、それまで青銅器や石に書かれていた文字が、竹、木片に書かれる様になったのも、文明の拡大を大きく促した。すなわち、より軽く、手に入りやすく、書きやすいマテリアルに文字を書くことで、文字のもう一つの重要な機能、コピーされ、伝播される機能が解放された。当然、これにより知識が多くの人間に伝播される。この様に、本来言語が持っているコミュニケーションの双方向性がWritingの機能として定着したこと、また情報をコピーとし伝搬できる媒体へと変化したことは、Writingの重要な2大転換点になった。阿辻さんの本から抜き出した漢字についてみると、この変化は表記法自体の変化を伴わずに起こっており、ほかの言語と比べた時、本当に奇跡的に思える。一方、他の言語では、この転換点でWriting自身が大きく変革している。
表子音文字と重商経済
実際全てのWritingで、双方向性の獲得、コピーによる伝搬可能性の獲得は起こっており、この変革が表記法の変化につながっている。例えば、日本語の万葉仮名に近い表音節文字から、ウガリット文字、そしてフェニキア文字の様に、表子音文字へと変化する過程は、農業を中心にした重農的帝国が中心だった古代世界で、地域間の政治・経済的な交流が高まり、商業、貿易を中心とする都市国家が生まれるのと一致しているように思える。
この文明や政治構造の転換がWritingの変革と相互作用しながら進むことをわかってもらうには、フェニキアとアルファベットの起源フェニキア文字ほどいい例はない。フェニキアが栄えた時代には、大規模な灌漑など、大きな権力を必要とする農業を基礎とした、原則的には周りとの交流の少なかった重農的政治体制間でも、相互依存的な政治的/経済的交流が盛んになっていた。この交流の時代には、一方的に何かを伝えるためにだけ使われ、その維持に多くの書記が必要だった第1期のWritingから、より双方向的で機動的な第2期のWritingへの変革が必要とされた。その意味で、海洋貿易に長けた重商国家フェニキア人はまさにこの変革の一翼を担ったと言っていい。

図2 レバノンの南の都市ティルスにあるフェニキア時代の遺跡(凱旋門)。
出典:Wikipedia
フェニキア人は海の人として有名だが、これが生き生きと描かれているのが旧約聖書の中でユダヤ人のバビロン捕囚時代について書かれたエゼキエル書だ。この中でフェニキアの都市ティルスの住民は次のように書かれている。
「海の出入り口を支配し、多くの島々を巡り、諸国の民と取り引きを行うティルス(図2)に向かって言いなさい。主なる神はこう言われる。ティルスよ、お前は言う。『わたしの姿は美しさの極み』と。お前の国境は海の真ん中にある。お前を築いた者は、お前の美しさを完全にした。彼らはセニルの檜でお前の外板を造りレバノンの杉で、帆柱を立てた。バシャンの樫の木で、櫂を造りキティムの島々の糸杉に象牙をはめこみ甲板を造った。美しく織ったエジプトの麻がお前の帆となり、旗となりエリシャの海岸から得た紫と赤紫の毛織物が甲板の日よけとなった。」(日本聖書協会 旧約聖書より)
太字をたどってもらうと、フェニキア人が多くの国と海上貿易を行ったことがわかると思う。エゼキエル書に挙げられている国だけでも、フェニキアの都市ティルスは、北シリア、ダマスカス、イスラエル、エジプト、アラビア、バビロニア、アッシリア、メソポタミア北部、アルメニア、中央小アジア、イオニア、キプロス、ギリシャ、そしてスペインまで活動範囲があり、ほぼあらゆる商品を扱っていたと書かれている。

図3:フェニキアの軍船: オールを2層に配することで、単位面積あたりの駆動力を挙げている。
エゼキエル書では結局ティルスもネブカドネザルにより征服される話で終わるが、しかしこれだけ多くの国々と交易を行う為には、双方に理解される表記方法が必須だったことは間違いない。実際フェニキアの貿易相手の国々では、表音節文字から表子音文字への転換が進み、それぞれの言語の音が表記できるようになっていた。この結果、言語が異なっても、双方向的な契約書などを交わすことが可能になったのだと考えられる。また以前議論したように、表子音文字は、アルファベットと比べても使用する文字が少ないことから、契約などをコンパクトに記録出来る。個人的感想だが、まさに商人にうってつけの表記法だ。このように、重商主義経済を基礎に形成された国家の発展には、各国家の相互交流の進展とともに、それに適応したWritingの変革が必要だった。
これと並行して、平等な市民の数が増え、Writingがより多くの人に使われることになったのも重要だ。ティルスも王が支配する都市国家で、重商主義といっても、多くの国と平等の交易を守るためには、組織化された軍隊を維持し(図3)、貨幣を管理する必要があり(図4)、王を中心とする国家体制は当然だったと思う。しかし、海洋を中心とした貿易で国家を成立させるためには、それに従事するかなりの数の平等な市民階級必要だったはずだ。このより広い階層の市民が使えるという要求が、フェニキア文字をはじめとする表子音文字の発明を促した。

図4 フェニキアの植民地で使われていた貨幣(Wikipediaより)
文明—Civilizationを支えるCivita(コミュニティー)の誕生とアルファベット
表子音文字であるフェニキア文字は、おそらく一人のギリシャ人によりギリシャ語のアルファベットへと作り変えられるが、これにはギリシャというフェニキアを超えた市民を中心にした新しい文明の誕生が大きな要因になったと思う。

図5:古代ギリシャについて全面的に参考にした書籍2冊
まず、私の古代ギリシャに関する知識の全ては、塩野七生さんの『ギリシャ人の物語』とOxford出版から出ているPaul Catledge著『Ancient Greece: A very short introduction』から出ていることを断って、話を進める。この2冊の本はともに、「ギリシャという明確な国はなく、ただギリシャ語を話し、文化を共有するギリシャ人がいただけだ」という説明から始まる。それを象徴するのがオリンピックで、ギリシャ人の統一のシンボルとして続けられた。近代オリンピックは戦争で中止になったことがあるが、『ギリシャ人の物語』によると、古代ギリシャのオリンピックは、村や都市の間で戦争が起こっていたとしても、オリンピック期間になるとその間休戦が成立して競技が行われたらしい。すなわち、ギリシャとはギリシャ語を話す様々なコミュニティーの集まりで、このような集まりから、アルファベットが生まれ、ヨーロッパの原点とも言える文明が花開き、これを私たちはホメロスの言葉を用いてヘレニズムと定義している。このように、文明:Civilizationという言葉がコミュニティー(Civita)の形成という意味から作られたとすると、この言葉は私たちの文明がギリシャから始まることを意味していることを主張しているのかもしれない。
要するに、エジプトやメソポタミアなどの大国のプレッシャーが低下した紀元前10世紀ごろに、同じ言語と文化を共有した、しかし互いに独立したコミュニティーがギリシャ各地で生まれ、それが集まってギリシャを形成していたと言える。それぞれのコミュニティーは独自の政治体制をとっていたが、かなりの数の身分の平等な市民が存在していた点が最も重要で、それ以前にはなかった文明の原動力になった。
『ギリシャ人の物語』によると、スパルタでは、市民権を持つスパルタ人1万、職人や商人のペリオイコイ7万人、そして農奴や小作農に当たるヘロット16万という人口構成になっていたようだ。一方アテネは、時代ごとに異なるが、商人や職人も含めたもっと多くの人口が市民権を得ており、市民権を持つ男性3万人とその家族で、市民が12万人もいたようだ。この市民の数を軍人に限定したスパルタでさえ、1万人という、原則平等に意思決定に関わる市民の存在は、Writingの大衆化を促進する。その時最も重要なのが、誰もが簡単に学べるという条件だったと思える。以前述べたように、表子音文字の場合、子音に続く母音は文字を見ながら頭の中で適当なものを選ぶ必要がある。この場合、一つの言語の中で発声の仕方に多様性があると(方言)、様々な誤解の原因になる。一方、アルファベットのように、子音と母音を組み合わせて、音を完全にコピーできるようになると、この様な間違いは消失し、また言語自体の標準語化も促進する。
以前述べた様に、ギリシャでは叙事詩に見られる様に、口述が重視され、そのためのレトリックが発展していた。これ自身、もちろんギリシャ語の標準語化に大きく貢献していたと思うが、しかし口述がいかに不正確で、効率が悪いかは考えるまでもない。ギリシャでは、多くの都市が4年に1回オリンピックで集まっていたが、この原則平等な市民の集会は、体育だけでなく、叙事詩などの文化も当然共有していたと考えられる。おそらく、このギリシャ語文化の共有も大きな力として、アルファベットの発明を後押しした。この様に、皆が話している音をそのまま表記できて、誰もが簡単に学べるアルファベットの発明を促す条件がギリシャには備わっていた。
この結果、ギリシャ語を表記するアルファベットがアテネで発明され、ギリシャの市民社会全体にすぐ浸透して行く。当時の市民階級での識字率の高さは、陶片追放に残された文字で知ることができる。紀元508年、クレイステネスにより、繰り返す有力者の闘争を収める為、アテネにとって害になる可能性のある人物を一定期間排除することを市民が投票で決めることができることを定めた「陶片追放」は、アテネ市民の全てに許された一種の投票権だが、これには追放する人間の名前を書く必要があった。

図6:追放する人の名前が書かれた陶片
最初法律が定めれられた時、10年間の追放を決議するには6000票を超える投票が必要とされていたが、これはすなわち万を越すアテネ市民がアルファベットを書くことができたことを意味している。
そして、新しく生まれたアルファベットを用いてまずホメロスの叙事詩が羊皮紙に書き写されたとされているが、もしそうなら、ギリシャのアルファベットは最初から豊かな文化をもたらすために発明されたと言ってもいい。書かれた内容のコピーには羊皮紙、パピルスなどが主に使われた。そして、コピーがコピーされるという形で、同じ情報が多くの人に届けられる様になった。
もちろんこのコピーされる過程の記録は残っていないが、今年7月、ホメロスのオデッセイの一部を書き写した粘土板が、オリンピアのゼウス宮殿で発見されたというニュースが飛び込んできた (BBCニュース参照:https://www.bbc.com/news/world-europe-44779492)。この粘土板は紀元後に書かれたものとされているが、羊皮紙、パピルスだけでなく、一般の市民でも手に入れやすい材料を用いて書き写されたことがよくわかる。
私見だが、アルファベット誕生はギリシャが到達した民主主義が結実した結果ではないだろうか。そしてアルファベットがヨーロッパ文化の起源となり得たのは、表記法としてのアルファベットの普遍性のみならず、平等な市民社会を支えることができるWritingこそが豊かな文化の条件であることを示唆している。そしてその結果、私が知っているだけでも、文学ではサッフォー(詩人)、ソフォクレス(悲劇)、アリストファネス(喜劇)、歴史家ではヘロドトスやツキジデス、そして哲学になると名前をあげてもきりがないが、最後には大御所プラトンとアリストテレスが控えている。
ヨーロッパはまちがいなくギリシャに始まる。
以上、フェニキアと、古代ギリシャを例に、Writingと文明の関係を考えてみた。あと、年末までは、Writingとバーチャルメディアについて続けて考えていく。
[ 西川 伸一 ]
2018年12月3日
あらゆる言語の音を、母音に対する文字と、子音に対する文字の組み合わせで表現するアルファベットの誕生は、支配階級に独占されていたWritingの民主化、大衆化をすすめるフェニキア文字から始まった進化の到達点だった。だからこそ、メソポタミアやエジプトでは多くの絵や彫刻が残されていたWritingを管理するテクノクラート=書記の姿は、ギリシャ彫刻にはもはや見ることができない。もちろんそれでも、Writingを習う市民と、そうでない人たちの階層は存在したが、Writing/Readingの大衆化へのパンドラの箱が開いて、文字を使う人の数が拡大した。
アリストテレスはその『形而上学』の冒頭で、「全ての人間は、生まれつき知る事を欲する」と述べているが、アルファベットが発明された後のWritingの歴史を見ると、まさにこの言葉が実感される。すなわち、人間の知りたいという欲求が、文字を使う人が一定の割合を超えることで解放され、Writingを用いて表現された個人のコンテンツを中心にした市民の文化が誕生する。
前回紹介したように、このWritingの大衆化と市民の文化はまずギリシャでその幕が切って落とされる。それまでギリシャの原点として口述されていたホメロスの叙事詩「イリアス」「オデッセイ」が一つの定本として確立する。それをきっかけに、叙事詩のみならず、悲劇、喜劇、歴史、そして哲学が市民の間で花開く。市民が文字を使ってコンテンツを作り、市民がそれを読む時代が始まった。これを可能にする新しい文字なしに、ギリシャ市民は生きることが出来なかったはずだ。
最初、階層的な社会で、上から下への一方的な通達に使うためのWritingが、同じ身分の市民の間のコミュニケーションの手段としてギリシャ市民に定着し、商業や産業だけでなく、娯楽や教養の核として、生活のあらゆる場面で必要になり、文化を牽引する。この文字に頼りきったようにみえるギリシャの文化状況については、これを憂いたソクラテスのぼやきからよくわかる。プラトンが書いたソクラテスとパイドロスとの対話、『パイドロス』から、天邪鬼ソクラテスのぼやきを聞いてみよう。

図1 岩波書店から出版されているプラトンの『パイドロス』。藤沢令夫訳
今回使ったのはこの本からの引用で、エジプト文字を発明したテウトに対して、全体に君臨していた神神タモスが応えた一節になっている。
「人々がこの文字というものを学ぶと、記憶力の訓練がなおざりにされるため、その人たちの魂の中には、忘れっぽい性質が植え付けられることだろうから。それは他でもない、彼らは、書いたものを信頼して、ものを思い出すのに、自分以外のものに彫りつけられたしるしによって外から思い出すようになり、自分で自分の力によって内から思い出す事をしないようになるからである。………。あなたがこれ(文字)を学ぶ人たちに与える知恵というのは、知恵の外見であって、真実の知恵ではない。すなわち、彼らはあなたのおかげで、親しく教えを受けなくてももの知りになるため、多くの場合本当は何も知らないでいながら、見かけだけはひじょうな博識かであると思われるようになるだろう」
たしかに文字についての本質をついている面もあるので、なるほどと思ってしまう。後に述べるが、キリスト教が支配し、聖書という「本の中の本」が文化を支配し、自ら考えることが抑制された長い中世の停滞を見ると、ソクラテスの指摘は的中していたのかもしれない。しかしギリシャ・ローマの市民文化の繁栄を見ると、Writingが大衆化したことによる効果は絶大で、市民はソクラテスが指摘したように知ったかぶりを繰り返しながらも、知識は拡大し、その後のヨーロッパ、いや世界の礎となる市民が中心になる新しい文化が誕生したことは間違いない。しかし、わざわざソクラテスがこのような警告を発しているのをみると、Writing/Readingへのギリシャ市民の依存性はかなりのものだったのだろう。
Writingの大衆化による文明の繁栄はそのまま古代ローマにも引き継がれていく。皆さんにもなじみのローマの文化を代表するよく知られた名前だけでも数えきれない。特に驚くのは、有名な歴史家プルタークの『英雄伝』をあげるまでもなく、軍人で政治家であったシーザー自身が『ガリア戦記』を著していることで、文字に残すということが、専門家以外でも日常化していたことがわかる。他に、文学や哲学で見ればキケロ、ヴェルギリウス、セネカなど、その後のヨーロッパの教養として学び続けられた名前が溢れている。
これだけコンテンツが創造され続けたギリシャ/ローマの文化は必然的に書物と図書館の発展を促した。エリク・ド・グロリエの『書物の歴史』(大塚幸男訳 白水社)によると、ギリシャではすでに本作りが産業化していたようで、出版社と共にそれを支える様々な職人、例えば写本を受け持つ職人がうまれ、さらに彼らの組合まで存在したらしい。ローマ帝国時代にはいると、ヨーロッパ全土からの需要に応えて、出版社は本の大規模な輸出まで行っていた。ホラティウスやキケロのような売れっ子の作家は決まった出版社がついており、著作料が払われていたのも現在と同じだ。逆に書物が思想を伝える力は圧倒的で、その結果当時から書物の内容を検閲するのは当然のことで、発禁になることもしばしばだった。このように、アルファベットの発明が開いたパンドラの箱は、文字使用の大衆化、そして全く新しい出版文化を誕生させる。
ただ、出版のためのコピーは全て写本だったため、原本を定めそれを忠実に写本して頒布するシステムが必要で、国家的もこの事業を後押ししていた。その中心になるのが何十万部もの蔵書を誇っていた図書館だった。ただ、残念ながらそこに集められていた本は完全に失われており、その内容を現在では知るよしもない。こうして、ギリシャ・ローマ時代に確立した出版技術はそのまま、中世の写本文化として受け継がれていく。しかしせっかく花開いた、Writing/Readingの土台の上に可能になった、出版という個人のコンテンツのコピー文化はその後ヨーロッパでは急速に下火になっていく。

図2 アレキサンドリアの図書館(出典:Wikipedia)
このギリシャ時代から続いてきた市民・個人を中心にするギリシャ・ローマ文化に対抗して、多様性のない、統一されたコンテンツを伝えることを重視する文化が、キリスト教のローマ国教化をきっかけに始まる。すなわち世界宗教キリスト教を通したWritingの市民への浸透だ。
ローマ時代にエルサレムから始まったキリスト教は、最初はユダヤ教の一セクトとして見られていたはずだ。しかし教義は世界宗教としての普遍性を有していたため、徐々にユダヤ教から独立し、世界宗教としてローマ帝国全体にひろがる。このキリスト教の世界的広がりについては、福音書と共に新約聖書の中心を占めるパウロの書簡が、イタリアから、ギリシャ、小アジア(アナトリア半島)、そしてイスラエルまで、ローマ帝国全土に拡がっていたことを見れば明らかだ。(これだけ長い書簡を各地に送るシステムが出来上がっていることは、一般の人間が羊皮紙などを用いた手紙のやりとりを普通に行っていた事を意味し、Writing がこの時代に市民の日常に深く入り込んでいることがわかる)。
君主制のローマでは、「この世の王」を主として認めないキリスト教は厳しい迫害をけることになるが、それでもキリスト教の拡大を止めることは出来なかった。この間、イエスの行動について述べた福音書と、パウロの世界の信者に当てた書簡は、それぞれのグループでコピーされ教会のアイデンティティーを守るのに大きな役割を演じたと思う。このとき使われていたのは、主にギリシャ語で書かれていた福音書や書簡、及びユダヤ教の教典としての旧約聖書だった。個人的見解だが、文字に記された教義なしに、ばらばらに離れた信者たちが迫害の中で統一した宗教を守ることは不可能だっただろう。このような世界宗教では、統一された書かれた規範の重要性は、エジプトやメソポタミアの王政よりはるかに高かったはずだ。このことは、多様なコンテンツを大事にするギリシャ・ローマ文化とは全く逆の、変わらない規範、すなわちコンテンツの統一性を重視する文化が新たに広がりを見せてきたことを意味する。もともとWritingは権力者がトップダウンで何かを一方的に伝えるために始まった(スマフォを用いて文字による会話が始まるまで、Writing自体はどんな状況で使われようと、著者から読者へと言う一方的な性格を持っている)。この意味で、新たに始まったキリスト教のWriting/Reading文化は、過去に後戻りしたことを意味している。もちろん、キリスト教の場合、おなじ方向を目指していてもこの世の権力とは無関係だった。
この変わらない教義を記すためにキリスト教徒が用いたのは、経済的理由で高価なパピルスでなく、羊皮紙だった。図2の棚に収められた巻物からわかるように、ギリシャ・ローマでの出版は、パピルスの巻物に写していくのが最もポピュラーだった。しかしパピルスは高価で、文字が読める市民だからといって、誰もが本を買って読める訳ではなかった。そこで、比較的安価に手に入り、削って何度も利用する事が出来、保管も優しかった羊皮紙がキリスト教では使われた。その後、書物の生産がキリスト教に収束する過程で、パピルスから羊皮紙へと、記録媒体も変化することになる。
迫害を受けながらもローマ帝国各地に散らばって信者をふやしてきたキリスト教は、ローマ帝国の強力な文化と同化する中で、普遍的世界宗教として313年には公認され、その後392年にはついにローマ帝国の国教となり、その後のヨーロッパを精神面で支配することになる。この結果アタナシウス派の教義以外の全てのキリスト教セクトは異端として、国家権力により禁止される。驚くのは、キリスト教内の異端宗派のみならず、本来のギリシャ・ローマの多神教も異教として禁止される。すなわち、キリスト教がキリストの死後400年間守ってきた、誰もが文字を通して学べる、多様性を認めない一つの真理という思想が、国家の思想として権力を得る。
この統一化した教義を制定するため、国教化により当然のように行われたのが、ウルガータとよばれるラテン語訳された標準聖書の制定だ。この作業を主導したのが、教皇の命をうけたヒエロニムスで、これにより旧約、新約が合わさった標準聖書の原型が出来上がり、一冊の「本の中の本」がその後のヨーロッパを支配するようになる。ヒエロニムスがヨーロッパ絵画の最も重要な題材の一人として、中世から千年以上にわたって何度も取り上げられてきたことは、このラテン語訳標準聖書の編纂がその後のキリスト教にとっていかに重要な事であったかを物語っている。図3にはルネッサンスの画家フィリッポ・リッピ (Filippo・Lippi)の描いたヒエロニムスの肖像を掲載したが、ダヴィンチからゴヤまで本当に多くの画家がヒエロニムスをとりあげている。

図3:標準ラテン訳聖書を編纂したヒエロニムス(ルネッサンスの画家 Filippo・Lippi作)
ヒエロニムスは、ヨーロッパ絵画の題材として最もよく取り上げられた人物。ルネッサンス以来ほとんどの画家が題材にしているのではないだろうか。
本来一神教は他の宗教に対して非寛容であるのが当然だが、統一された教義の枠にはめて世界をとらえることが、キリスト教の国教化で唯一正統な思想となり、ルネッサンスまでのその後のヨーロッパ文化を完全に決定してしまう。くり返すがこの思想は、個人により創造される多様なコンテンツをベースにしたギリシャ・ローマの文化とは相容れない。そのため、ギリシャ・ローマの市民文化は排除され、多くはイスラム圏に移って維持されることになる。そして当然のように、個人のコンテンツに基づく出版文化は、これを契機に消滅していく。グロリエの書物の歴史によると、この結果本を扱う商業を頂点に成立していた出版産業は完全に崩壊したらしい。また、公的私的に維持されていた図書館も、閉鎖され、略奪され、せっかく生まれたこの出版文化の成果も完全に消滅してしまう。その結果、ヨーロッパの大学や、都市で、教会から独立した世俗の活動が盛んになる12世紀ごろまで、出版文化は教会の中に閉じ込められ、独自の発展をとげる。
聖書のヨハネによる福音書は「始めに言葉ありき」で始まるが、キリスト教の歴史を見ると、権力とは全く無関係の一般人が思想を共有するためには、何よりも「書かれた言葉」が必要であったことがよくわかる。逆に言うと、原始キリスト教以来守り続けられた「文字で書かれた言葉」が、実際の権力をもしのぐ強い力を持ちうることが証明された。だからこそ、ギリシャ・ローマの出版文化を完全に否定したキリスト教も、標準聖書を誤謬なしに写して多くの人に届け、そこに書かれたメッセージを伝えるため、独特の出版技術を発展させることになる。その究極が中世の最大の芸術の一つと言っていい、絵入り写本だった。
[ 西川 伸一 ]
2018年12月17日
ギリシャ・ローマの市民文化を支えた出版業は、ローマ帝国によるキリスト教国教化により、一度完全に下火になる。結果、出版という事業は教会や修道院でのみ行われるようになり、この状態が中世まで続くことになる。しかし、書物を造る出版技術という面では、中世は出版技術にとってはきわめて重要な時期で、このことはヨーロッパ各地に残る地域の文化を生かした多様な絵入り写本に見ることが出来る。図1にアイルランドで8世紀に制作された有名なケルズの書を示す。美しいだけでなく、地域性や個人の創意に満ちた絵入写本の聖書を見ていると、キリスト教の許容力に驚嘆する。このような写本に現れる多様性をみると、神が万物を絶対的に支配する一神教の思想の枠の中で、神を人間に近づけ、それによって個人の自由を認めることが出来た、カソリックの特徴を見て取ることが出来る。

図1 ケルズの書 アイルランドで8世紀に制作された聖書の写本。ケルト模様で装飾された美しい本で、キリスト教という題材の中で、それぞれの地域の才能が生き生きと作品に向かったことがわかる。
キリスト教の世俗性は教会に飾られている像や絵を見れば一目瞭然だ。キリスト教のルーツであるユダヤ教では、モーゼや預言者が描かれても神が絵に描かれることはない。イスラムもそうだが、モスクやシナゴーグの中は、幾何学的な装飾が施されて美しいが、決して人の姿を描いた絵が飾られてはいない(少なくとも私が見た限り)。しかし、カソリック教会の内部には多くの人間の像が描かれ飾られている(図2)

図2 ユダヤ教 シナゴーグ、カソリック教会、イスラム教モスクの内部の写真。(Wikipediaより)
これは、キリスト教が世界宗教としてローマ帝国の国教になる過程で、あえて神と人間の区別を曖昧にする教義を正当化していったからではないだろうか。例えば、復活したイエスの神性を巡っては、国教化の過程で「三位一体」を正統と定め、イエスより上に絶対的神がいると考えるアリウス派などは排除される。そして、三位一体の当然の帰結として、マリアは神の母であるとする「テオトコス」と呼ばれる重要な考え方が、431年エフェソスの宗教会議で正統として認められる。この考えは、現在でもロシアを含む東ヨーロッパの教会の多くに残り、崇拝の対象になっている。この考えを表現するため、聖母マリア(テオトコス)が神を抱く聖母子像と、マリアの死の床にキリストが降臨してマリアの霊を取り上げる「Domination of Theotokos」の像がセットになっていることが多い。

図3 テオトコスとDomination of Theotokosの教会画(出典:wikipedia)
テオトコスやDomination of theotokosがキリスト教神学的にどう考えられているのか、私は全く不勉強だが、マリア=人間が神を生んだ事を認めることは、人間と神を対称化させようとしているように私には見える。例えば仏教では、人間は成仏できるとしている。すなわち人間が仏になれるわけで、人間と仏が対称化されていることを意味する。この結果世界中の仏教寺院では人間の形をした仏像が必須の崇拝対象としておかれており、何の違和感もない。カソリック教会でもこれと同じで、人間の像が崇拝対象として設置されているように思う。もともと、ローマやギリシャは偶像崇拝が当たり前の国だったが、私にはキリスト教がローマの国教になる過程で、一種の偶像崇拝を潜在的に容認してしまったのではないかと思う。実際、マリア崇拝はキリスト教で当たり前のことと考えられている。ただ私自身は、このキリスト教の世俗性が、カソリックが統一的に支配する中世でも、個人の能力生かした芸術をはぐくみ、潤いをあたえ、多様な文化を生み出す原動力になったと思っている。

図4 ヒエロニムス・ボッシュの三連祭壇画「快楽の園」の一部。地獄を書いているが、実際には中世信じられていた多くの逸話が想像力豊かに書かれている。(出典:Wikipedia)
ヨーロッパの美術館に行くと、最初の部屋は中世アートの絵画や彫刻が展示室であることが多いが、ビザンチンからロマネスクにかけての絵を見ていると、中世の絵画がいかに多様だったのかがよくわかる。題材は全てキリスト教でも、作品は多様性と創意に満ちている。すなわち、一定のルールの枠内で創造的な文化が花開き、多様性に満ちている。
同じことは、写本ではもっと顕著に表れている。図1で示したケルズの書の写本でも、統一した思想の枠のなかで、個人や民族(この場合ケルト民族)の伝統と創意が溢れているのがわかる。描かれた時代は中世ではないが、図に示したヒエロニムス・ボッシュの地獄絵を見ると、その創造力に驚嘆するはずだ。私自身は、ボッシュの絵はヨーロッパ中世絵画の到達点だと思っている。
このように、中世の美術や工芸の面白さはこのカソリックという統一された枠の中にもかかわらず、個人の創意と工夫が解放され、統一されたテーマと緊張感を持っている点だと思う。この最適の例が絵入り写本に描かれた細密画だろう。図5は中世に蓄積された絵入り写本の頂点の1つと言っていい、当時の「売れっ子」細密画家、ジャン・フーケの絵が描かれている写本『エティエンヌ・シュヴァリエの時祷書』を示すが(図5,左側)、書物に描かれた絵画としての頂点に到達していると言える。同じ図の右側は、以前脳の記憶を説明するときに使った、アントワープ美術館にあるフーケの絵画だが、この絵の不思議なグラフィック的感性も、フーケが写本の細密画で売れっ子だったことを知るとうなずける。

図5:左はジャンフーケが絵入り写本に描いた細密画。右は、アントワープ王立美術館に飾られているムーランの聖母子像(出典:Wikipedia)
印刷の発明と新しい市民文化
このように、「物を知りたい」という欲求は、結局中世でも抑えきれることはなかった。最初、教会や修道院の中に閉じ込められていたこの欲求は、教会とは別の世俗の力が増すに応じて、都市や大学などに分散し、ガスのように溜まり続けていた。実際、12世紀になると修道院での写本は下火になり、主に大学で写本が行われるようになる。この結果、キリスト教とは無関係の歴史や小説といった世俗の題材も出版の対象として選ばれるようになっていた。一度教会に吸収された出版業に関わる職人組合も新しく生まれ、出版と販売数は増加し続けていた。そしてこのガスが大爆発したのが、グーテンベルグによる活版印刷技術の発明だ。
活版印刷というと、ドイツ・マインツのグーテンベルグの名前が最初に上がる。しかし、印刷自体はグーテンベルグの印刷法の発明よりはるか以前から、漢字圏では木版印刷による書物がつくられており、わが国でも8世紀には印刷による書物が作られている。また活字を使う活版印刷もグーテンベルグより200年前に、朝鮮で始まったことが知られている。そして、出版文化は、中国でも仏典や四書・五経などの宗教・道徳書だけでなく、文学や詩を中心に個人のコンテンツを尊ぶ文化にも大きく寄与している。ただ、漢字は文字としてはきわめて複雑なため保存できる活字として必要な数を常に用意することは難しく、朝鮮の活版印刷はほとんど普及する事はなかった。その結果、漢字圏での印刷は、木版に刻み込んだ版木を用いるスタイルから変化することはなく、常に修業を積んだ優れた版木職人を必要とした。版木による印刷の1つの頂点を見ることが出来るのが、ユネスコの世界遺産に選ばれている韓国の海印寺に保存されている八万枚にも及ぶ仏典の版木だろう。この版木から印刷された仏典が我が国にも運ばれ保存されているが、この完成に職人が何人必要だったのだろうか?

図6 韓国海印寺に保存されている大蔵経の版木のコレクション
活版印刷は発展しなかったが、それでも漢字圏でこれほど早くから印刷文化が花開いたのは、なんと言っても東漢での紙の発明だろう(桑原 隲蔵 「紙の歴史」による。写真はWikipedia)。
パピルスのように、草の線維をそのまま並べてシート状に仕上げるのではなく、紙は線維を完全に分離して、それを漉いてつくる。そのため、セルロースさえ分解できれば、どんな植物からでもつくることが出来、唐時代に紙の製法が8世紀にイスラム圏に伝わると、パピルスはまたたくまに紙に置き換えられる。この紙が11世紀ヨーロッパに導入されると、ヨーロッパで木版印刷も始まり、あとはグーテンベルグによる1450年の印刷術の完成を待つだけになる。

図7 グーテンベルグと彼の発明。グーテンベルグの肖像、活字、プレス機、そして有名な 行聖書 (出典:Wikipedia)
グーテンベルグについては多くの本が書かれており私自身が今回最も参考にしたJohn Man著『The Gutenberg Revolution』(Penguin Books)によれば、彼は若いときから書物を印刷するという大きな構想を抱いて、技術を開発し、事業のための資本を集め、最終的に目的を実現した、今で言う起業家だった。この準備段階で、彼は金属細工師として生計を立てながら、
を始め、必要な技術の全てを開発している。まさにイノベーターと言っていいだろう。
この事業を進めるため、彼は大きな借金を抱えることになる。この借金を返すためには、最も売れる本を最初に印刷する必要があった。この要求に最もマッチしたのが、図7に示した42行聖書で、カソリック教会の金銭的支援も受け、この聖書は当時のベストセラーになった。ただ、それでも借金を全て返すには及ばなかったようで、出資者だったフストにより、彼が開発した技術のほぼ全ては借金のかたに没収される。さらに悪いことに、この技術を共に開発してきた右腕シェッファー(フストの親戚でもある)もフストとともにグーテンベルグの工房を離れ新しい印刷工房を作ることになったため、グーテンベルグより資金力の豊かな競争相手がうまれる。その結果印刷出版という点では、シェッファーとフストの事業は拡大し、現在にまで至る印刷事業の原型は彼らにより完成される。
ちょっとグーテンベルグの悲劇を強調しすぎたかもしれないが、彼が事業家として成功したかどうかよりはるかに重要な点は、出版業が特殊な技能を必要としない職人により運営できる、「儲かる商売」になった事だ。木版印刷と比べると、活版を作るスピードは速く、しかも職人の熟練機関は遙かに短くて済んだ。その結果、経済的波及効果は大きく、例えば、Quarterly Journal of Economics(126: 1133, 2011)に掲載されたDittmarの論文によると、1450年にマインツ一箇所だった印刷所が、50年後には205都市に広がっていたことが示されている。当時のヨーロッパで都市の数は1811だったので、なんと11%の都市で印刷業が行われていたことになる。しかも、印刷業が存在した都市はそれ以外と比べた時高い成長率を示しており、印刷業がそれ自身として地域経済に寄与するだけでなく、産業全体に大きな影響を持っていたことが分かる。この結果、樺山紘一著『本の歴史』(ふくろうの本)によると、1500年までの50年間で驚くなかれ250万冊の本が出版されたと推定されている。これは中世全体で出版された本の数に匹敵する数で、アルファベットの発明により大衆化を成し遂げたものの、まだWritingに内在していたが限界が、印刷の発明により取り除かれ、新しい情報革命が起こったと考えられる。
この限界とは何だったのか、次回からWriting の情報としての側面を見ていくことで、新しい情報革命について考えてみたい。
[ 西川 伸一 ]
2019年1月7日
前回グーテンベルグの印刷術の発明が、言語およびWritingに内在していた限界を取り除くことで、人類の文明に全く新しい可能性を開いたことを述べた。振り返ってみると、言語誕生の後解放された人類の脳が持つ高いコミュニケーションポテンシャルは、文字を中心に様々な技術開発により高められてきた。その意味で、印刷術が可能にした飛躍は文字誕生に匹敵するのではないかと思える。そしてこの飛躍を実感できるのが、ルネッサンスの文明復興ではないだろうか。
いつルネッサンスが始まったかは私にとってはどうでもいい。おそらくルネッサンスが始まるきっかけはローマカソリックの政治/文化支配力の低下だったことは間違いがない。すでに見てきたように、ギリシャではアルファベットの発明をきっかけに、市民が文化を担う文化の大衆化が始まった。この動きは、ローマでも受け継がれる。しかしキリスト教がローマの国教になった頃から、市民が文化の担い手になるというルートは閉ざされ、文化の大衆化はキリスト教の大衆化と同義になった。もともとキリスト教は、世界宗教として大衆化という方向性を強く持っていたが、自由な市民を情報としての出し手とする許容力は当然皆無だ。宗教としては当然のことだ。その結果、出し手が教会に集約した上で文化の大衆化が進む。当然文化を担った書籍/記録は全て教会や修道院に閉ざされ、写本による書籍の生産技術も修道院に集中する。
しかし、カロッサの屈辱に象徴される教会の権威も、その後世俗の力に侵食され始める。政治的には、この変化を王権の強化や、商業を中心とした都市の力が高まってきたことに見ることができる。ルネッサンス人文主義の立役者メディチ家が、金融業でフィレンツエに君臨したのはその典型例だ。文化という面で言うと、もちろんメディチ家に限らず、修道院以外に王侯や都市の支援を受けた大学などが情報の出し手として機能し始め、写本による出版もこのような拠点に移って行った。このように14−15世紀にかけて始まるルネッサンスの前に、情報の発信源が多様化し、市民の手に戻り始めていたと言える。
この大きなルネッサンスというトレンドの中で、印刷術の発明という情報革命が起こるが、フランスやドイツなど北ヨーロッパ、イギリスへのルネッサンスの広がりは、まさにこの印刷術の発明および普及と軌を一にしている。逆にいうと、ルネッサンスで始まった文化復興の動きを抑制していた重要な要因が、印刷術により取り除かれたことになる。全くの私見だが、この要因とは、言語や文字だけでは不可能だったコミュニケーションの大衆化阻んでいた様々な要因、大衆化したコミュニケーションの同時化を阻んでいた要因、そして科学を阻んでいた要因だったと思っている。この3要因は、印刷術そのもので解決したものではなく、印刷術がもたらす可能性を最大化する過程で解決され、新しい文明復興に大きく寄与した。
情報の大衆化
まず印刷術は、ルネッサンスから始まりつつあった情報の受け手の量と質を大きく変化させ、大衆化した。印刷術が開発されても、勿論書籍は庶民には高嶺の花だったに違いない。しかしOur World Dataと言うサイト(https://ourworldindata.org/books)に掲載されている統計表を見ると、オランダでの本の値段は、印刷術が行われる前と後で8割安くなっている。価格が安くなるというのは、もちろん生産コストが下がったことも意味するが、それだけでなく価格を下げても儲けが出るほど需要が旺盛だったことを意味する。実際、ルネッサンスの人文主義思想により、例えばコシモディメディチが市民に開かれた図書館を建て、さらにルターも図書館の重要性を強調したように、本は多くの読者と共有するべきだとする思想も広がりを見せていた。従って、印刷術により安価になった本は、個人、公を問わず多くのライブリーのレパートリーを豊富にしたことは間違いない。
このように、本に対する強い需要があったおかげで、それを商売にする出版社(印刷所)がヨーロッパ中に設立される。図1はWikipediaから転載したヨーロッパの印刷所の分布を示しているが、最初マインツだけではじまった印刷業が、1500年にはヨーロッパ中に広がっているのがよくわかる。また印刷業の分布をみれば、情報の出し手としての教会の独占が崩れたこともよくわかる。
図1:15世紀のヨーロッパの印刷所の分布 1450年から50年でこれほどの広がりを見せたのは驚きだ。(出典:Wikipedia)
実際、印刷技術によって、文化の正の大サイクルが回り始めた。情報にアクセスできるようになった受け手が多様化し、その中から新たな出し手が生まれ、それがさらなる受け手の多様化を生むというサイクルだ。中でも象徴的なのは、1300年代に完成したダンテの「神曲」が、グーテンベルグの印刷術発明後20年には印刷され出版されていることだろう。すなわち、売れるコンテンツを求める印刷業が急速に拡大を初めていた。今も昔も、人間は常に新しい情報を求めるようにできている。
出版の同時性
印刷業によって取り除かれたもう一つのコミュニケーションに対する障害要因は、本の制作にかかる時間だろう。もともと人間のコミュニケーションはジェスチャーであれ、音節言語であれ、リアルタイムで起こる過程だった。ところが、文字の発明により、出し手のコンテンツを、長い期間繰り返して伝えることができるようになった。しかしWritingによるコミュニケーションは、まず伝えたいコンテンツを書き起こすために長い時間を要する。例えば、石に彫ったり、粘土板やパピルスに書き残すには、専門職による作業が必要で、エジプトやメソポタミアではこれが書記の役割だった。そのため自ずとコンテンツの内容は限られる。
その後ギリシャ・ローマの市民文化の発展により、写本が盛んになり、様々なコンテンツを伝える体制は整ったが、コンテンツが長ければ長いほど、それを伝える本の製作には時間がかかったことは間違いがない。簡単になったアルファベットだったとしても、ホメロスの叙事詩をパピルスに書き留めるのに、どれほどの時間が必要だったのだろう?しかもそれをコピーして、もっと多くの読者に伝えるためには、さらに長い時間がかかる。おそらく、著者の存命中に、著書についてのコミュニケーションが図れる可能性はほとんどなかったように思う。
印刷術も、活版を作る過程は写本と同じ時間がかかるが、その後のコピーを作るという点ではその時間はほとんど0に近くなったと言っていいだろう。このおかげで15世紀からすでに、著者の生存中にコンテンツを出版して、広い範囲に流通させることが可能になった。もちろん今と比べるとはるかに長い時間を要したと思うが、これにより批判や批評といった双方向性のコミュニケーションが、出版物を通してはじめて可能になった。これが、17世紀の科学誕生にも重要な役割を演じることになる。

図2雑誌German Historyに2006年発表されたWeber
の論文。
このコミュニケーションの同時性を求める市民が生まれ、それに答えようと印刷業が努力する過程は新聞の歴史をみるとよくわかる。ブレーメン大学のWeberの論文「Strassburg, 1605: The origins of the newspaper in Europe」によると(図2)、現代的な意味で定期刊行(週刊)新聞は、出版社のオーナーで、執筆者でもあったJohann Carolusにより、1605年にストラスブルグで発刊された「Relation Aller Fuernemmen und gedenckwuerdigen Historien」が最初だった(図3)。驚くのは、その後1620年までには、ヨーロッパ各国で同じような定期刊行物が発行されていたことで、出し手も受け手も、リアルタイムのコンテンツを求めいたことが良くわかる。

図3:世界初の週刊新聞
もちろんこのような週刊の刊行物が読者を得るためには、印刷術だけでなく、新聞を運ぶ郵便システム、そして主に政治について様々な意見を述べる評論家が育っている必要があった。このため毎週読者に届ける定期刊行物が生まれるまで、グーテンベルグの発明から150年要している。しかし、定期的ではないが「かわら版」のような印刷物だと、かなり以前から発行されており、ルネッサンスの市民たちが情報を貪欲に求めていたことがわかる。
この点で、私が最も注目したいのは、ドイツの人文主義者Sebastian Brantが1492に発行したFluigblatt(ビラ、チラシ)だ。図4に示したチラシには、アルザス地方のエンシスハイムに落下した隕石について、Brantによる説明が絵とともに書かれている。これが現代まで残っているということは、かなりの数印刷されたのではないだろうか。図4下に示すように、隕石は100kgを越す大きさで、大気圏に突入した爆発音は大変なものだったと思える。当然その原因について多くの人が心配したはずだ。これに対して、このようなチラシを発行し、その原因を知らせ、安心させる作業が、グーテンベルグの発明以来50年も経たないうちに行われていたのには感心する。

図4 Sebastian Brantがエンシスハイムに落下した隕石について記載したチラシ(Flugblatt)と、その時落下した隕石。(Wikipediaより)。
さらに重要なのは、Brantのチラシは、まさに科学的観察についての報告書と言っていい点だ。もちろん定期刊行物の主要な目的は政治について出来事や意見を掲載することだったが、新聞を可能にする条件はそのまま、現象についての意見を述べ、批判を受けるという科学を可能にする条件に合致している。
そこで次回は、科学の誕生と印刷について考えたいと思う。
こうしてみてくると、印刷術という技術革新によって起こった変化は、PCからスマートフォンを技術革新として個人のレベルでおこっている情報革命に大変よく似ていることがわかるのではないだろうか。要するに文字が誕生して以来、人間はコミュニケーションの大衆化、同時化を常に望み続けてきており、最初の糸口が印刷術の誕生だったと言える。そして今、一人の出し手が瞬時に1億人の人に簡単に情報を届ける時代が来た。それが何をもたらすのか、Writingの最後に考えてみたい。
[ 西川 伸一 ]
2019年1月15日
近代科学の誕生と印刷術
印刷術の発明と普及は、17世紀近代科学の誕生のための必須条件だったと思っている。そこで印刷術により新たに抑制要因が取り除かれた最後の領域として、近代科学について考えてみよう。その前に、近代科学誕生とはなんだったのか、ざっとおさらいしてみたい。近代科学誕生の条件の詳細については、このコラムの延長として計画している「生命科学から読む哲学書」で詳しく扱おうと思っているが、今回は全ての詳細を省略して近代科学誕生についての私の考えを短くまとめる。従って、これから書くことは全て私の独断と偏見に基づくと思っておいてほしい。
17世紀近代科学を誕生以前
「科学とは何?」と聞かれると、「世界で起こる現象を理解するため、一定の手続きに基づいて進める探求」と答えている。理解したい対象はこの世界で起こった、あるいは起こっている現象なので、人間個人や社会での出来事も当然含んでいる。ただ、「一定の手続きに基づいて」という但し書きが重要で、それに基づかない限り科学は成立しない。この手続きこそが17世紀の近代科学誕生で明らかにされたことだ。
世界で起こる現象を理解し、説明することはもちろん古代から行われた。この探求の最初のピークは、古代ギリシャに見られる。タレスに始まり、アナクシマンドロス、デモクリトス、ピタゴラス、ユークリッド、アルキメデスなどなど、自然の探求を目指した人たちの名前は枚挙に遑がない。その後ローマ時代に入ってこの動きは低調になるが、これは情報の出してがキリスト教に集約したためで、「神の業」についてあれこれ探求すること自身がナンセンスな話ということになってしまった結果だ。しかし、一神教の影響がなかった古代ギリシャにおいても、近代科学成立に必要な重要な条件が欠落していた。
これが「作り話を拒否する」という科学の条件だ。例えばギリシャ科学の集大成と言えるアリストテレスは、世界の現象は4つの原因による結果だとする「4因説」を提案したことで有名だ。すなわち、世界で起こる現象は、1)質料因、2)作用因、3)形相因、そして4)目的因(最終因)、のいずれかの原因で起こるとした。確かに、現象には必ず変化するものが存在し、それが何からできているか(質料因)、何がそれに働きかけているか(作用因)などは、現象の原因として的確に把握できる。また無理をすれば、それぞれの物が本来あるべき場所があるとする形相因も、たとえば比重、さらに極端に言えば、一般相対性理論での時空の曲がりで説明できるかもしれない(勝手な妄想なのであまり気にせず読み飛ばして欲しい)。しかし、最終因・目的因となると、それが何かよくわからない。まず物理学的には目的を持ち出すことはタブーだ。一方、形相因や目的因についてのアリストテレスの説明を読むと、このような原因の存在を最初から決めてかかっていることがわかり、説明自身は作り話としか思えない。
この傾向はキリスト教世界に入るとさらに深まる。天地創造やノアの箱舟など疑ってはならないドグマをもとに現象を説明しようとするため、作り話がなんの抵抗もなく無限に導入され、それを疑うことも許されないという状況が続くことになる。科学にとって、この状況が変わったのが17世紀で、宗教からの介入を棚上げにして、作り話を排除する体制が徐々に整備される。この時の最も重要な立役者がデカルトとガリレオだ。
17世紀近代科学誕生
図1 マリアの死に際してキリストが現れ、マリアの霊を取り上げる、マリアは神の毋とする考え方を示している。特に東欧の教会には数多くの同じパターンの絵画が飾られている(筆者撮影)
なぜ西欧より科学が進んでいた、中国やイスラム圏で近代科学革命がおこならなったのかについては、いつか議論したいと思っているが、一つの重要な要因として、キリスト教(特にローマカソリック)が、世俗的な一神教だったことが大きいように思う。イスラム教やユダヤ教徒比べたとき、マリア(人間)がキリスト(神)の母になれるという教義は(図1)、ある意味で人間と神が比較的対称的な関係を持つことを許した考えと捉えられる。その結果、カソリックやギリシャ正教の教会には、仏教寺院や多くの多神教の寺院と同じように人間の像が溢れることとなっている。にもかかわらず、一神教の立場から、自然現象を「作り話」を駆使して説明することが行われており、例えば天地創造やノアの箱舟といった話に見られるように、全ての作り話が唯一の神に集約していた。その意味で、うまく神の世界を現実の世界から切り離すことができれば、科学的にも、政治的にも作り話に対する対処がしやすかったのだと思う。言い換えると、世俗的一神教では、神が人間に近いおかげで、精神的に神の世界を現実から棚上げすることが17世紀になって進んでも抵抗が少なかったのではないだろうか。またルネッサンスの古代ギリシャ文明の再評価も、この動きを後押ししたように思う。一方、イスラム教のような神と人間が絶対的に非対称な宗教では、神が起源となる作り話を拒否することは、まず不可能だろう。
ただ、これだけでは不十分だ。あとで述べるように、西欧で印刷業が急速に広まり、コミュニケーションの大衆化、同時化が進んだことが、17世紀西欧で最初に近代科学が誕生した重要なきっかけになったと思う。
デカルトと近代科学
デカルトの考えの詳しい解説はまたの機会に譲って、ここでは彼の近代科学誕生に果たした役割についてだけ述べる。

図2 ルネ・デカルトの肖像画と1637年に出版された方法序説
デカルトといえば「我思うゆえに我あり」という有名な言葉が書かれている「方法序説」だが、この本はなんと当時で3000部印刷されたと何処かで読んだことがある。この本は哲学の本なので、現在でも3000部というのはかなり売れた部類にあると言えるのではないだろうか。実際、デカルトはカソリックに睨まれていたものの、当時の売れっ子思想家だった。(出典:Wikipedia)
デカルトの近代科学誕生への貢献は、主観主義と2元論に尽きると思う。まず主観主義だが、方法序説を読むと、彼自身の経験からスカラ哲学など当時の権威を疑い続けて最終的に、疑っている自分から始めるしかないことに気づく過程が書かれている。すなわち「我思うゆえに我あり」だ。この自分から始め、自分で考えるというのは科学の最も重要な条件であることは誰も異論がないだろう。ただ、神の言葉(=教会の教義)が絶対であった当時、これを明確に語たることは簡単でなかったはずだ。
もう一つの貢献が2元論で、これはその後合理主義の指針として現在まで続いている。個人的には、教会との一種の妥協のようなアイデアとして出てきたように思えるが、「心と身体」を神の領域と人間の領域に分離した点で、その後の科学の発展に多大な影響を及ぼした。すなわち、心のように本質的に理解できないことがあり、これを無理に説明しようとすると作り話になる。説明がもともと難しい領域は、神の領域として棚上げして、理解可能と思われる身体の領域に集中すればいいという考えだ。極端にいうと「わからないことは放っておけばよく、まずわかることから取り組め」ということだ。当たり前の話だが、これをはっきりさせたおかげで、わざわざ全てを説明しなくても、理解できることに集中すればいいことになり、この考え方が近代科学を推進し、現代まで続いているように思う。現代、演繹主義とか、機械論として批判されるのは、全てこの2元論の話だ。
ガリレオ・ガリレイと近代科学
ガリレオというとみなさんは「それでも地球が回っている」と地動説をとなえ、当時のカソリックの天動説を否定した科学者を思い出すと思う。もちろんそうなのだが、図3に示した彼の本、「偽金鑑識官」を読むと、彼が「作り話」を拒否し、第三者と合意を図るための方法、すなわち科学の方法を明確に示した人であることがわかる。デカルトが示したように、自分で考え、わからないことは後回しにするにしても、「思う我」が到達した結論が他の人の同意を得られるかどうかはわからない。残念ながらデカルトの著書でもこの問題は全く触れられず、彼の提案するテーゼも基本的には同意されるものとして提示されている。
これに対しガリレオは、それがどれほど正しい概念でも、他人に同意を強要し、押し付けることを科学は拒否すべきだと語る。そして、実験や数学のように、第三者と共通の概念を共有するための手続きだけが、同意を形成するための唯一の手段であると主張する。そして返す刀で、教会関係者がなんの根拠もない思いつきを正しいと他人に押し付けていることは、科学でも哲学でもないと強く主張した。その意味で、この「偽金鑑識官」を読んでみれば、ルネッサンスで世俗に侵食され力が衰えていたとはいえ、当時のカソリック教会がガリレオの思想を許すはずはないことが理解される。

図3 ガリレオの肖像と1623年に出版された偽金検査官
(出典:Wikipedia)
読者の皆さんもご存知のように、ガリレオは数多くの観察や実験を行ったことで知られている。ピサの斜塔での落下実験は最も有名だが、多くの実験道具を開発している。また、望遠鏡を用いた天体観察を行い、木星の衛星の発見など、実際大きな研究上の業績もあげている。しかし、17世紀近代科学誕生に関していえば、「贋金鑑識官」に込めた彼の思想が、最も大きな貢献をしたと言える。
近代科学の誕生と印刷
デカルト、ガリレオにより、近代科学の過程が、
ことで、作り話を排除し、合意された概念を積み重ねる過程として確立する。言い換えると、科学者が、第三者と同じ手続きを認め合い、概念について合意を得ることを最も重視する人として明確に定義された。そして、この原則は現在も何も変わることはない。
ただ、この過程を実現するためには、科学者個人が方法を理解し、実験や数学的な検討を行うだけでは不十分だ。すなわち、着想し、検証した概念を、複数の第三者に見せて批判を仰ぎ、問題があれば直した後、共有できる概念として再度確立し直すという、今でいう論文を発表するための査読システムに似た過程が必要になる。すなわち、出版物を通したコミュニケーションが必須になる。これを可能にしたのが、印刷術で、近代科学誕生にグーテンベルグの貢献は大きい。
古代ギリシャでも、自然現象を説明しようとした人たちの中には、同じように第三者と概念を共有したいと考えた人たちは必ずいたと思う。ただ、口述にしても、自分でパピルスに書き下ろすにしても、まとめた考えを多くの人に見てもらうためには、写本を繰り返すしかなかった。ところが、これにはどうしても時間がかかるため、ある程度の同時性が必要な批判しあい、合意を撮る過程が実際には成立できなかった。キラ星のように様々な思想が生まれた古代ギリシャを読んだ時、どうしても個人が勝手に思いつきを語っているという印象を持ってしまうのはこれが原因だと思う。
おそらく観察や数理の適用という点では、古代ギリシャもかなり優れていたと思うが、近代科学が定立するためには、科学に必要な批判し合う関係を維持するために必須の、印刷術と出版業がまだできていなかった。一方、近代科学誕生前の16世紀のルネッサンスには、科学についての本の出版がすでに行われていた。例えば有名なコペルニクスの「天球の回転について」は1543年に出版されている。すなわち、グーテンベルグの発明から100年後には、なんと科学についての本が出版され、販売されていたことは驚きだ。近代科学を誕生させる、インフラストラクチャーは16世紀に準備が進んでいたと言っていいだろう。
最後に、本稿での最初の質問に戻ろう。「科学とは何か?」
これに対して最も有名なのが、「反証する方法がないものは科学ではない」としたカール・ポパーの答えだが、証明も、反証も大衆的、同時的コミュニケーションなしにありえない。その意味で、近代科学は印刷術に支えられて始まったと言っていいだろう。
[ 西川 伸一 ]
2017年6月1日
今回から言語の起源と進化について見ていこうと思っている。おそらくこのコーナーを読んでいただいている読者の多くの方は、「え!言語の起源」と驚かれるのではと思う。
「進化研究を覗く」というタイトルを見て生物進化について知りたいとこのコーナーを訪れていただいている読者のほとんどは、純粋生物学分野の人たちのはずだ。ところが神経系の進化についての話が終わった頃から、「意識、自己」と、話が進化研究から脱線を始め、フロイトの話まで進んでしまった。もうたくさんと密かに思いながら、ここまで付き合ってきていただいたのではと本当に感謝している。それでも、私の専門とは全く異なる言語まで進まないことには「進化研究を覗く」は完成しないと思っている。というのも、言語もまた38億年生命進化の結果だからだ。
一見進化とは無関係な話がなぜ延々続くことになったのか振り返ってみると、進化を「環境によるゲノム情報の選択」ではなく、「環境による統合された生物情報の選択」として捉えるために、生物個体に集合している情報を一つ一つ検討し始めたことに始まっている(図1)。
図1:人間には様々な情報がかたまって存在している。説明文中
進化を環境と生物情報全体の相互作用として捉えることの重要性は、動物進化を考えるとよくわかる。例えば神経記憶という情報がゲノム情報に加わると、環境が大きく変化して個体の適応性が低下しても、生存にかなった場所を見つけて生き延びることができる。すなわち、ゲノムに直接選択圧力がかかるのを、他の情報で防ぐことができる。隕石の衝突のようなビッグインパクトを除けば、この戦略は種の多様化に大きく貢献している。従って、閉じられた試験管内の大腸菌の進化で見られるルールを、自由に動くことが可能な動物に当てはめることはできない。
こう考えて、ゲノム、エピゲノム、神経回路などを、生物個体に集合している情報という観点からひとつひとつ検討してきた。地球上に存在する全ての生物情報媒体が集まっているのは図1に示したように人間だけだが、ゲノム、エピゲノム、神経回路は生物進化の異なる時期に現れ、その最後に人間だけに言語が発生している。そして、その都度生物多様性は急速に増大している。おさらいの意味で、もう一度これらの情報としての特徴を説明しよう(図1)。
生命誕生前の地球は完全に物理学の法則だけに支配され、情報として認められる因果性は存在しなかった。そこに生命誕生と並行して地球上最初の情報、すなわち塩基配列をコードとして用いたゲノム情報が誕生する。最初は機能を持つ高分子として現れたRNA(or DNA)が、たまたま4種類の塩基でできていたため、記号性を持つことになり、最終的に生命の情報媒体へと発展することになる。またこのおかげで複製が可能になり、情報の指示による化学反応の制御が可能になった。おそらく、ゲノム情報誕生が先で、そのあと現在のような生物が誕生したと思われるが、ゲノム情報は「非物理的因果性」として物理化学的因果性と合わさって、それ以前にはなかった生物という新しい因果システムが地球上に誕生し、生命の進化が始まった。
最初生物にはゲノムしか情報媒体は存在せず、当然進化は、核酸が本来持つ傾向からうまれる多様性を、環境が選択するという単純な図式の下で進む。組み替えによるゲノム情報の直接交換も行われることはあるが、原則として同じ情報が同時に個体間で共有されることはなく、環境にフィットした個体が選ばれ、子孫を残すことで進化が進む。
個体間で外界に関する解釈を共有できないというゲノム情報の難点は、クロマチン構造の調節を介して遺伝子のon/offを調節する機構、エピジェネティックス機構が誕生することで解決される(例えば温度の違いによりクロマチン構造は書き換えられる)。この機構は最初、情報が複雑化し大きくなったDNAをコンパクトにまとめるために発達したと考えられる。その後このクロマチン構造に関わる分子を遺伝子on/offを指令する情報として利用し、細胞分裂を繰り返してもそれを維持できるエピジェネティック機構が誕生すると、環境の状態を特定のクロマチン情報に転換して記憶し、異なる細胞同士で外界の刺激について同じ解釈を共有し対応することが可能になった。
具体的には、例えば環境により同じクロマチン構造の変化が異なる個体に誘導されることで、環境変化に合わせて遺伝子発現を変えることが可能になり、ゲノムの違いに影響されずに、個体を様々な環境で維持することができるようになった。この様々な環境に個体として適応できる能力により、さらに生物の多様化が可能になる。こうして生まれた同じクロマチン構造を共有するという特徴は、ゲノムは変化しないまま細胞が分化することを可能にし、生殖細胞と体細胞が区別される多細胞体制が可能になり、生命がさらに多様化する原動力になった。
しかし完全な情報という観点から見るとエピゲノムにはいくつかの問題がある。例えば、クロマチン構造による環境の解釈は、個体間・細胞間で共有できても、原則として子孫には伝わらない。すなわち、環境への対応は、個体が生きている時一回きりになる。また、ヒストンなどの分子の化学的変化に基づく反応は早くない。そしてなによりも、この情報はゲノムの存在が必須で、副次的な情報でしかない。
このエピゲノムが持つ情報としての独立性の欠陥を解決したのが、次に誕生した新しい情報媒体、神経ネットワークだ。もちろん個々の神経細胞自体には情報媒体としての機能はない。原始的な神経に見られる様に、感覚器がそのままシグナルを運動器に伝える様な構造では、外界の記憶が発生したとしても、それは全て神経細胞自体のクロマチン構造の変化が直接反映されたものだ。しかし、いったん神経細胞同士が結合してネットワークを形成すると、細胞自体の変化を超える、回路の構造が媒体となる情報が発生する。すなわちネットワーク結合のパターンと、ネットワークを構成する個々の神経細胞の特性の変化が統合された、DNAからは完全に独立した情報媒体が成立する。
この結果、外界の変化に迅速に反応するとともに、外界の変化を記憶したり、その変化に応じて行動を起こすことが可能になった。また、神経細胞興奮という同じ原理の上に形成される回路自体を複雑化することができるため、感覚、運動、記憶などのそれぞれの回路を複雑に連結することで、厖大な外界からの刺激の中から一部のみを選択するための基準、すなわち身体とは独立した神経回路の自己を形成し、また内外からの刺激を取り込んで、この神経回路の自己を常にアップデートすることが可能になり、さらにこの自己を基準として厖大な外界の刺激の中から重要なシグナルだけを選んで神経ネットワークの自己を書き換える過程、すなわち意識を生み出すことに成功した。
この神経ネットワークの機能について、道で知り合いに出会って、挨拶を交わすという行動を例に考えてみよう。人と出会うと、視覚、聴覚、あるいは嗅覚から特定の刺激を受けることになる。刺激自体は物理量の変化だが、全て神経ネットワークに転換した上で処理されるため、まず感覚神経の興奮へと転換され、神経ネットワークに入ってくる。刺激が神経回路上に表象される過程で、物理化学的刺激は完全にシンボル化された情報になる。この内外の刺激により誘導された興奮は、すでに形成されているその時点の神経ネットワーク(=神経ネットワークの自己)を基準に選ばれ、この選択フィルターをパスした興奮だけが、その個体の既存の神経ネットワークと相互作用を始める。フィルタリングされる段階で、視覚、聴覚、嗅覚などの入り口の異なる情報が統合されるが、人に出会うセッティングでは、入力刺激の統合過程で形とや色が備わった顔のイメージが形成され、場合によっては音や匂いが統合される。こうしてできたイメージは、次にネットワークの自己を形成する様々なサブネットワークと結合することで、刺激の内容が特定されると同時に新しいイメージに発展する。このイメージに基づいて、挨拶を行うという選択が行われ「○○さんご無沙汰しています。××大学での実験はうまくいっていますか」と実際に挨拶することになる。
もちろん神経ネットワークの形成や活動は、ゲノムやエピゲノムの支配を受けているが、基本的には神経興奮という同じ原理を共有する神経細胞のネットワークに表象され、様々なネットワークを自由に重ねたり、連結することで、独自の情報を形成していくのが神経ネットワークの特徴だ。情報としての特徴を見ると、個体レベルで環境変化に対する反応を起こせるエピゲノム情報と比べても、外界の変化に対する反応は極めて早い。もちろん外界についての解釈を、他の個体と同時に共有することも可能だ。しかし、こうして生まれた回路パターンや活性は、まだその個体一回きりで、世代を超えて子孫に伝えることは難しい。
迅速に情報を処理できる神経ネットワークの進化へのインパクトは大きい。例えば急に襲ってきた様々な変化に対して、迅速に対応し、過去の記憶もたどりながら広い範囲を移動して新しい生存環境を探す可能性が生まれ、より広い環境の変化に集団で対応することが可能になった。
図2:環境が限られると、自然選択によって生物多様性は減じると考えられる(上段)。しかし、自分のゲノムにフィットする新しい環境を求めて移動できれば、当然個体の多様化に応じて種も多様化できる(下段)。
図2に示す様に、ダーウィン進化論では多様化した個体の中で、環境に最もフィットする個体が子孫を残すと考えるが、このままでは集団内の多様性は減少する(オレンジ色ばかりになる)。例えば中立説では、この問題を選ばれた個体のゲノムのほとんどは選択に貢献せず、そのまま多様性として維持されると考えることで、自然選択により多様性が増大することを説明していた。エピゲノム機構がこれに加わると、個体レベルでの適応が起こり、ゲノムに対して直接の選択圧が加わることを軽減することができるようになるが、結果的には中立説の考えの少し異なるバージョンといってもいいかもしれない。いずれにせよ、中立変異やエピゲノムの効果が合わさり、ゲノムの多様性を維持したまま個体数が維持され、集団内のゲノム多様性は自然選択で減少することはない。
この上に神経ネットワークが誕生すると(文字通りの動物の誕生)、ゲノムから独立した情報に基づいて個体自体が、自分にフィットした環境へと移動することで、種の多様性を増大させることができる様になった(図2下段)。このように、エピゲノムや、神経情報も統合された生命情報全体と環境との相互作用を考えることが進化を考える上でいかに重要かを再確認いただけただろうか。
しかし図1に示した様に、エピゲノムも神経情報も身体から独立していない、その個体一回限りの情報だ。このため、極めて稀な例を除いて、その情報が子孫に伝わることはない。この限界を初めて破った生物情報が言語だ。この情報の誕生が、進化にどれほどの影響を及ぼしたのかは、現在100億人を越す人間が、地球上の隅々に生きていることを見るだけで理解できると思う。すなわち、科学をはじめこの人間の発展を支えた人間にしか存在しない高次機能は全て言語誕生から始まっていると言っていい。情報としてみると、同時代の個体と同じ情報を共有し、さらに一部を子孫に伝えることができる点が大きな変化だったと言える。
この様に進化研究を知るためには、他の動物とは際立って異なる人間の進化を抜きに終わることはできない。したがって「進化研究を覗く」も、図1に示した様に、この人間特有の進化の原動力となった身体から独立した情報、言語・文字・バーチャルメディアにいたるまで、その誕生と、生物進化へのインパクトについて考えるつもりにしている。しかし、ゲノム誕生、エピゲノム誕生、神経ネットワーク誕生と比べると、この作業はさらに困難であることは書いている私にもよくわかる。
というのも、これまで見てきた新しい情報の誕生は、少なくともそれに対応する物理化学的過程を構想することができた。例えば、RNAワールドで、偶然塩基配列がインデックス情報となり、最後にアミノ酸と結びついてシンボル情報になるといった過程だ。もちろん全ての過程を完全に解明できているわけではないが、エピゲノムも、神経ネットワークもそれが生命情報に統合される過程をなんとか説明できた。
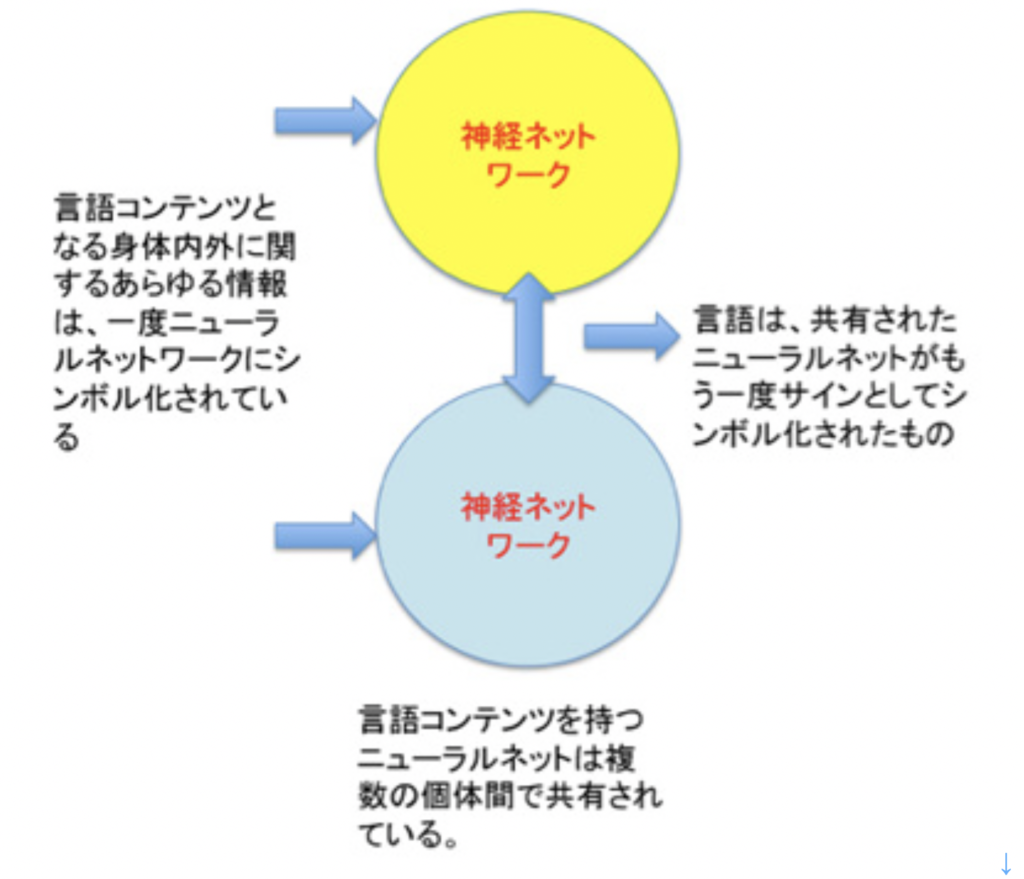
図3:言語情報の生物学的研究の困難
一方言語誕生は神経ネットワークの活動から生まれたもので、それに対応する物理学的過程自体も神経回路へとシンボル化しているため、物質的痕跡が全く存在しない。このことを図3に示した。繰り返すが、言語のコンテンツとなる様々な情報は、物理化学的に記述される現象に対応していても、言語として表現されるまでに、一度ニューラルネットに転換され表象されるため、シンボル化されている。この特定の内容が表象されたニューラルネットを複数の個体が共有することができると、今度はそれが再度シンボル化され、言語ができる。このように、シンボルの体系から新たなシンボルの体系が生まれることが言語の生物学的研究の最大の困難で、私自身も言語の起源について説得力のあるアイデアがあるわけではない。
結局思いつくまま、言語誕生に関わる様々な研究を紹介することになると思うが、今後も是非お付き合いいただきたい。
[ 西川 伸一 ]

図1 出典:wikipedia
言語学と生物学を結びつけたチョムスキー
写真は米国MITで50年以上にわたって言語学研究を続けているノーム・チョムスキー(Noam Chomsky)だ。本や論文は読んだことがなくても、名前を知らない人はほとんどいないと思う。
ギリシャ時代から現代まで、言語について多くが語られてきたが、近代言語学の父と呼ばれるソシュールも、一つの言語(例えばフランス語)の歴史や、言語の構造比較に基づいて、言語と文明・社会との関わりを考える研究が中心で(と言っても私は岩波書店一般言語学講義しか読んではいないが)、言語がどのように生まれてきたか、私たちの脳の持っている言語能力とはなにかについて述べることはなかった。
これに対し、チョムスキーは、まだ脳科学がほとんど発展していなかった1950-1970年、言語、特に単語を集めて文にする統語論について多くの著作を表し、統語過程の背景には、すべての言語に共通で普遍的な「生成文法」と、学習によって身につける「個別文法」が存在することを提唱した。そして、この生成文法は生まれつき備わっていると考えた。それまで、言語とは一人一人の個人が白紙の状態から学習するものと考えていた当時の言語学にとっては、この考えは大きなインパクトがあったはずだ。
1965年に書かれた「統辞理論の諸相」(福井直樹、辻子保子訳、岩波文庫)のなかで、生成文法について彼は次のように語っている。
「生成文法とは端的に言えば、明示的で明確に定義された方法を持って文に構造記述を付与する規則のシステムに他ならない。ある言語の話者が、誰でもその言語についての自分の知識を表している生成文法を身につけ、内在化していることは明らかである。だからと言って、話者がその文法の規則がどのようなものなのか気づいているとか、気づくことができるようになるとか、あるいは話者が自分の言語についての直接的知識を言語化したものが必ず正しいとか、そういったことにはならない」——すなわち、わたしたちが意識しないで、一定のレベルの意味のある言葉を話すことができるのは、私たちが生来身につけている生成文法による。
「生成文法は、話者が自分の言語知識について語るかもしれないことではなく、話者が実際に知っていることを明示的に述べようとするのである。」——すなわち、生成文法は私たちの頭の中で物語(文章)が形成されるプロセスに関わる。
「生成文法とは話者のモデルでも聴者のモデルでもないと念を押すことは、無意味ではないかもしれない」——すなわち、話者も聴者も同じ生成文法を共有している。
そしてこの生成文法の背景にある、母国語の種類にかかわらず人間の脳には全てに備わっている統語能力を普遍文法と呼んだ。今風に言えば、進化過程で変化してきた人間共通の脳神経回路構造の中には、外界、内界から得た表象をもとに物語を形成するための十分条件が備わっており、この人類共通の言語回路が、外からの学習によって書き換えられると、言語別、個別の文法が形成されるとする考えだ。
現在の脳科学に十分親しんだ読者の皆さんは、一定の統語能力が発生直後の脳回路に備わっていると考えることにはそれほど違和感はないはずだ。
もちろん、チョムスキーがこの革新的な説について述べた1950-1970年には、脳についての理解は限られていた。このため、個体の多様性が自然に発生することが自然の原理であることを説得するため、ダーウィンが家鳩などの多様化の例を執拗に提示したのと同じように、チョムスキーも生成文法や、普遍文法を、文例の分析からわかる現象論的には自明のこととして議論を展開した。しかし例えばガゼルの子供が生まれた時から歩けることは直接観察できるが、子供が私たちが十分理解できる言葉を話すようになるのに3年近くかかることを考えると、彼の説を脳科学的に検証するのは難しい。事実、チョムスキーがこの考えを最初に述べてからすでに50年が経っているが、生成文法に対応する脳神経回路を特定するような研究はまだないと言っていいだろう。
ただはっきりしているのは、チョムスキー以後、言語や言語能力の発生が重要な課題として多くの研究者に認められ、賛成・反対を問わず、生成文法や、普遍文法は必ず言語の発生に関する議論の中心になって来たことだ。この現象は一部の人には、チョムスキーがこの分野に神のごとく君臨しているように見えるらしい。普遍文法が神話になってしまった現状を憂いたトマセロ(サルを使って言語や社会性について優れた研究を展開している)は「Universal grammar is dead」というコメンタリーで、普遍文法から言語を考えるのではなく、バイアスなしに脳科学から言語の普遍性と多様性を考えることの重要性を述べている。私も全く同感で、脳科学的にも言語学的にも普遍文法という概念にまだエビデンスに基づいた内容が伴ったとは言い難い。
チョムスキーらの最近の論文
しかし、トマセロがこの批判的コメンタリーを書いた2009年から比べると、チョムスキー自身も変化しているのは当然だ。彼の今を知るには、最近書かれた論文や総説を読むのが一番だ。そう思いついて、この原稿を書くため彼が最近書いた総説を探してみた。彼は1928年生まれで、現在89歳に近いと思うが、驚くことに今年に入ってすでに単独、あるいは共著で論文を3編発表していることがわかった。そこで彼が現在生成文法についてどう考えているのか、これらの総説に探ってみようと考えた。これらの総説論文から、チョムスキーの考えの論点を整理し、今回言語の発生を考える時の課題をリストすることが今回の目的だ。

図2 今回紹介する2012年に書かれた総説
幾つか論文をダウンロードして読んだ中で、共著だが2012年にTrends in Cogitive Scienceに発表された論文は、図も多く、また最初から言語とチョムスキーの統語理論を脳科学的に考えようとしているのでこの論文を中心に紹介することにした(図2)。この論文は、普遍文法の脳科学を目指して書かれているように思える。この論文を紹介しながら、必要に応じて彼が今年単名で発表した論文(Neuroscience and biobehavioral reviewsに発表した論文)にも言及する予定だ。
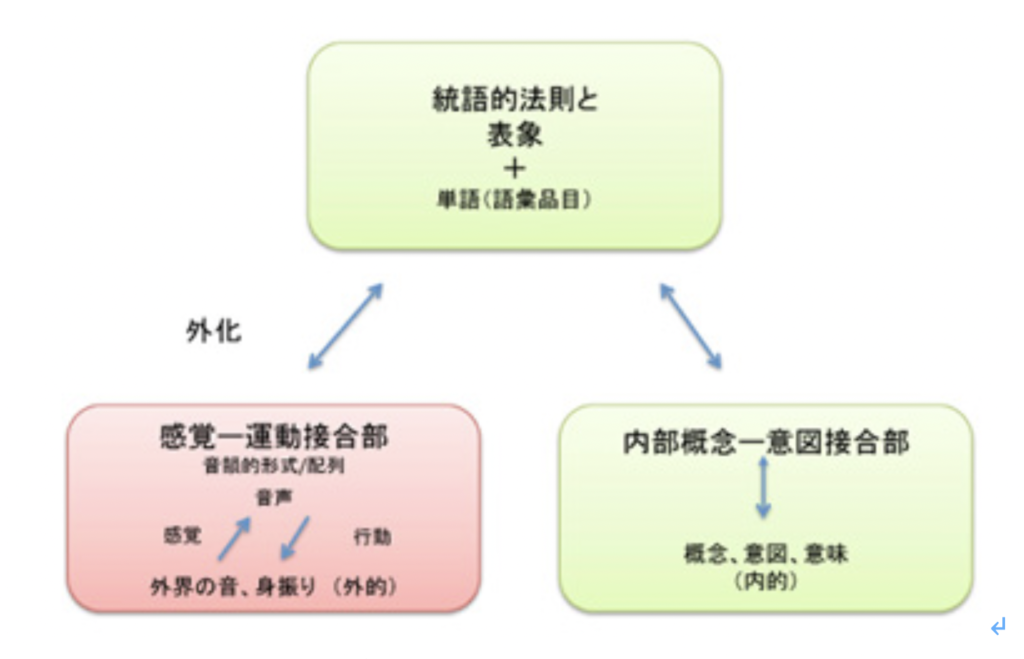
図3 総説の図1を改変して再掲している。内容は本文参照。
脳に普遍文法を探る
チョムスキーの最初の頃の著作と今回読んだ総説を比べると、50年経った今、彼が自分の理論を脳科学として位置付け直そうとしているのがよくわかる。総説に掲載された図を改変した図3は、脳内の言語回路と、他の機能を担う回路との関係を描いたものだが、言語回路が結合する最も重要な脳回路は、外界との関係に関わる感覚運動回路と、概念や意図の形成に関わる内的な高次機能回路で、言語回路とそれぞれとの接点を、1)感覚〜運動接合部、と2)概念〜意図接合部と呼んでいる。すなわち言語活動とは、脳内の言語回路が、外界との相互作用を行う感覚・運動系、および脳内で概念、意図、意味などを発生させる高次脳回路を媒介することで成立すると考えている。
例えば、何かを考えてそれを言葉に出す時、内的高次回路、言語回路、感覚運動回路という順番でプロセスが進み、意味のある文章が発話される。あるいは、友達に出会って話をする時などは、友達を認識する感覚運動回路からスタートするが、あとは三つの回路を行ったり来たりすることになる。
この脳内言語活動において普遍文法に相当するのは、進化と発生過程で生まれた、人間共通の基礎言語回路になるが、現段階でこれが言語に特化した回路だと考える必要はないだろう。要するに、言語を可能にする言語能力が新しい脳回路形成過程で人間のみに現れたと考えればいいと思う。
言語学の課題は、この言語回路で行われる過程の解明だが、最近の総説では脳内に散在する様々な表象を「merge」(混合)して、新しい表象(文章)を生み出すのが言語回路の役割だとしている。残念ながらこのmergeを行う脳内での計算が、ランダムではなく、意味を生み出す法則についてはこの総説でもわからないままだ。文章には単語が順序だって並んでいることから、単語を並べることがこの回路の仕事と考えてしまうが、決して単語を並べることがMergeではないことを強調している。すなわち、言語回路は表象を順番に並べるのではなく、mergeによって塊(句)にまとめることが主要な役割だと考えている。確かに文章が発話されるとき、単語が順序立てて連なっているが、頭の中の塊をそのまま同時に伝えることができないという物理的制限のせいでこうせざるを得ないだけのことだ。また、各言語で個別の文法が発展しているのは、おそらく様々な文化的背景のなかでコミュニケーションをより正確に進めることが要求されるからだと言える。
このmergeについて、2017年のチョムスキーの総説では、言語も脳のコンピューティングであることを強調し、脳の中ではあらゆる表象が脳回路の活動であることを強調している。これは私が前回言語発生研究の難しさとして述べた、神経系ではあらゆるものが電気回路にシンボル化されているということと同じことだ。即ち “eat” “apple”のようにeatとappleがmergeしてeat appleという新しい表象が生まれるとき、eatもappleも神経回路にシンボル化して存在している点だ。チョムスキーは1990以降、この過程で働く法則としてminimalist programを提唱し、この総説でもStrong minimalist thesisについて述べているが、統語の法則がニュートン力学と同じように法則として存在するとする考え方で、脳科学的に説明することが難しい概念だと思う。この法則性は別として、単語が脳回路でどのように表象されているかという点は、言語を考える上で極めて重要な課題で、のちに詳しく議論したいと考えている。
以上まとめると、普遍文法とは頭の中に異なる表象を集めて新しい意味をもつ表象の塊を作るために生まれてきた人間特有の能力で、この能力は脳内で物語を作るために進化したもので、決してコミュニケーションの必要性で進化したとは考えていない。ただこの能力は、2次的にコミュニケーションにも使われるようになる。この結果、学習が必要な個別文法がコミニュケーション言語には必要になった。このように言語能力が、コミュニケーションのために進化したと考えないのがチョムスキーの普遍文法の一つの柱だが、これについても多くの異論があり、特に、言語の原子とも言える単語がどのように成立するのか考えるためには、コミュニケーションの問題を抜きにしては語れない。
すでに述べたが、チョムスキーの総説では、具体的な対象に対応するシンボルとして単語が存在しているのではなく、具体的な対象や行為と関連付けられる脳内の表象が単語に対応する。ただ、具体的な事物が脳内で表象されるとき、他の表象ともともと結合して塊を作っている。例えばサルはappleを理解するときは常にeat appleとして表象しているらしい。この、具体的な事物や行動が脳内回路に表象されるときすでに様々な塊と様々な程度で連結しているという可能性は、言語発生を考えるとき極めて重要な鍵になると私は考えており、これも別に議論するつもりだ。
図3を確かめる実験
詳しくは述べないが、この総説の目的は、図3で示した脳内のデザインを、実際の脳に存在する回路と対応させることが可能であることを示すことだ。例えば、この総説を読んで、人工的文法(artificial grammar)を用いた新しい言語認識研究法があるのを知った。人工文法とは、自然には存在しない単語や句の並びを聞いたときの脳の反応を調べる研究だが、生成文法が提唱されて以来50年、急速に進展した脳のイメージングによりこのような実験が可能になった。
紹介されている実験を詳しく説明することは避けるが、このような方法論の進歩によって、言語を処理する際、脳内でどのネットワークが活性化されるのかがわかるようになってきた。言い換えると、言語理論と脳回路とを結びつける可能性が生まれたわけで、今後普遍文法など統語論の脳科学を進めるときには欠かせない手法になりつつある。もちろんチョムスキーも、この分野の進展を取り込み、理論を発展させようと考えている。
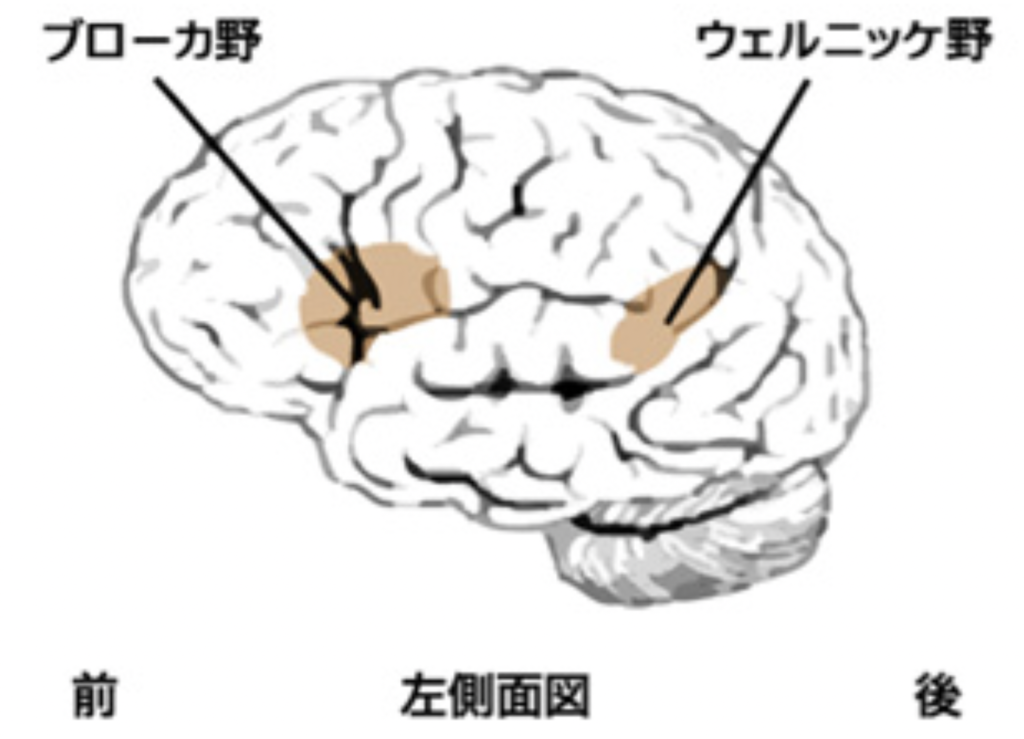
図4 ブローカ領域のウェルニッケ領域の脳内での位置。失語症の研究から、ブローカ領域に障害が起こると文法的に整った文章の発話が困難になる。一方ウェルニッケ領域が障害されると、言葉を聞いて理解するのが困難になる。ただ実際にはもっと複雑で、改めて紹介する。
(出典:wikipedia)
例えば、失語症の研究から文法的に整った文章を話すときに必須であることがあきらかにされていたブローカ領域(図4)は、運動性言語中枢と呼ばれ、図3で示す感覚運動回路を通した発話過程に関わっていることがわかっていた。人工的に単語を並べたセンテンスと、自然言語によるセンテンスを比べる研究から、ブロカ領域はどちらの刺激でも活性化されるが、自然言語を処理するときは、言語を聞いて理解するのに必要で知覚性運動中枢と呼ばれるウェルニッケ領域も同時に活動することなどがわかってきた。このような脳科学的データを集めることで、図3をより詳しい回路図へと仕上げることができるはずだ。
チョムスキーらも総説の最後に、言語過程には知覚性言語中枢(ウェルニッケ)と運動性言語中枢(ブロカ)に加えて、幾つかの脳領域が言語処理に関わっており、また単語や意味に関わる独立した領域も存在していることがわかってきた。これら領域のなかに具体的事物の表象がどう形成され、またそれがMergeされるのか、脳科学として研究が可能であることを強調している。この問題については、言語中枢の局在化の問題として取り上げて考えてみたい。
このように、新しいチョムスキーの総説を読むことで、脳科学として言語発生を扱うための課題を整理することができた。これ以外にも、
1)言語能力の進化と人類学、
2)チョムスキーは言語が進化するのではなく、言語能力が進化すると明快に断じているが、本当に言語自体は進化しないのか?
などについても、次回以降順次考えていきたいと思っている。
前回チョムスキーの考えを詳しく説明しないまま、チョムスキーから始めてしまったので、この分野に興味を持っていなかったみなさんにはわかりにくい話になったと思う。説明している私自身も、彼の初期の生成文法についての考えを完全に理解できているわけではないので、余計にわかりにくくした。しかし、彼は言語学を脳科学や進化学、すなわち生物学として捉えるべきだと最初に考えた人で、そのことを伝えられるだけでも意味があったと思っている。言語の問題は生物学・脳科学の問題であるとする彼の考えに私も100%賛同する。チョムスキーから始めた限りは、これから言語発達に関する様々な問題を、脳科学の問題として具体的に取り上げていこうと思っている。もちろん、議論のなかで必要とあればチョムスキーの考え方も参照しながら進めたい。
さて第一回は、「道具と言葉」というタイトルで、人間特有の脳機能の進化について考えてみたい。
現在地球上で言語を話すのは人間だけだ。そのため、人間と例えばチンパンジーの遺伝子を単純に比べることで、言語の遺伝子に到達できるという話をよく聞く。言語を考える時、わたしはこのような単純な発想は間違っていると思う。言語は、何か一つの能力が備わったことで急に現れたものではない。すなわち、長い進化の過程で様々な能力が蓄積した結果が、人間特有の性質の一つである言語に象徴されていると思う。一番わかりやすいのが複雑な発声を可能にする解剖構造だが、複雑な発声だけなら鳥類、あるいはイルカだって独特の解剖学的構造を発生させ実現している。一方、多くの点で人に近いと言える類人猿はこのような複雑な発声はほとんど不可能だ。このように、生物進化で獲得されてきた様々な独立した能力の蓄積が言語能力の背景にあるとするなら、言語に限らず人間を特徴付ける様々な高次機能を検討することも、言語の条件を知るためには重要だ。
例えば道具を使う能力について考えてみよう。この能力も、これまで人間特異的能力として考えられてきた。最近になって、サルやカラスなどが道具を使う能力を持つことが明らかになっているが(後述)、それでも道具が日常化して進化の道筋が変わったのは人間だけだろう。
世界で最も古い330万年前の石器は、アウストラオピテクスが居住していたと考えられるオルドワンで見つかっている。
図1 チンパンジーから現代人までの頭蓋、脳容積、歩行様式、歯、樹上生活の変化。石器が見つかるアウストラロピテクスから急速に犬歯が退化する。肉を道具で切り分ける社会が生まれ、個体間の関係も大きく変化した。その意味で、一夫一婦制も石器による起こった変化だとすら考えられる。(Nature Knowledge Project, Pontzer et al, Overview of hominin evokutionより引用:https://www.nature.com/scitable/knowledge/library/overview-of-hominin-evolution-89010983)
道具が生まれる前後の猿人から人間への変化を調べると、まず犬歯が消失する。これは道具を使うことで肉を引きちぎる必要がなくなり、口の中で噛み続けるためには犬歯が邪魔になったのだろうと考えられる。しかしもっと興味深いのは、男女の体格差が急速に減少することで、オーストラロピテクスでは2倍もあった大序の体格差が、エレクトスになると1.5倍と現代人の1.2倍に近づいている。おそらく、道具を使って肉を切り分けることで、1匹のオスがすべてを支配する権力構造が解消し、おそらく一夫一婦が原則となった共同体が生まれたことが、男女体重差の解消の背景にあるのだろう。
道具の研究が重要なのは、文字が出来るまでは歴史的検証に必要な遺物が全く存在しない言語の起源研究と異なり、石器という物証が残っていることだ。道具は当時の知的レベル、移動と交流など、当時の生活をかなりの確度で教えてくれる。
例えば道具の形態や出土の状況から、共同で狩りをしながら複数の家族が一緒に暮らしていることがわかれば、これを維持するためにどのレベルのコミュニケーションが必要かどうかわかるし、さらには一夫一婦制を前提とする社会なら、浮気のないシステムを守るためにかなり高度なルール設定が必要だったことも推察できる。このように、道具を通して明らかになる社会構造は、言語発生の条件について重要な資料になることまちがいない。
では道具を使う脳が発達するためにはどのような条件が必要だっただろうか?
これを知ろうと、道具を使う人間以外の動物の研究が進んでいる。この方向の研究のルーツは、有名な類人猿の生態研究者Jane Goodallが、葉っぱを使ってありを釣り上げるチンパンジーの報告に遡る。その後の研究の結果、様々な動物が道具を使うことが示されてきたが、中でも小枝を木の穴に挿入して虫をおびき出すニューカレドニアやハワイのカラスの行動は、最も原始的な道具使用の始まりとして研究が行われてきた。
この道具を使うハワイのカラス(アララと呼ばれている)の論文を読んだ時、不思議なことにチョムスキーの普遍文法を私は思い出してしまった。そこで、まずこのアララについての最新の研究から紹介しよう。
このアララが小枝を拾って、木の穴に突っ込んで隠れている虫をおびき出して食べる様子、あるいは小枝が落ちていない時は、枝を折って使う様子が、この論文を掲載したNatureによりYouTubeにアップされているので一度見て欲しい(https://www.youtube.com/watch?v=ZOUyrtWeW4Q)。
この論文では(Rutz et al, Discovery of species wide tool use in the Hawaiian crow、Nature, 537, 403, 2016 (http://www.nature.com/nature/journal/v537/n7620/abs/nature19103.html)),この小枝を使う行動がカラスに生まれつき備わった遺伝的なものか、あるいは学習により獲得されるものかを調べる目的で、7羽のカラスを道具を使う他の個体から隔離した環境でヒナから育て、教えなくても道具を使うようになるかを調べている。結論は明確で、全てのカラスが他の個体から習うことなく、道具を使えるようになるという結果だ。
「道具を使うという「文法」が生まれつき備わっており、学習する必要がない」と解釈しても良さそうなこの結論を読んだ時、私はすぐチョムスキーの普遍文法に似ているなと思った。しかし、枝を見た時、それをくわえて穴を探り、虫をおびき出すという一連の行動を実行する全過程が、ロボットのように生まれた時から頭に組み込まれているのだろうか?
論文を詳しく読むと、行動の順序が生まれた時から頭の中にプログラムされているのかどうかは判断が難しいことがわかる。最も重要な問題は、全てのカのカラスの道具使用を確認するのに5ヶ月もかかっている点だ。もし、行動の全てがプログラムされているなら、巣立ち(1ヶ月程度)直後から道具を使ってもよさそうだ。
ここからは私の想像だが、アララが習わないで道具を使えるのは、もともと習性として持っている遺伝的性質と、自分の数ヶ月にわたる学習が組み合わさった結果だと考えればどうだろう。もともと鳥は巣作りのために、木切れなどを拾って穴に突き刺す行動を習性として持っている可能性は高い(確かめたわけではない)。この習性を繰り返しているうち、たまたま虫をゲットする経験が続くと、この行動は記憶され、固定化し、枝を道具として虫取りに使うようになると考えることもできる。
このシナリオでは、他の個体の行動を学習する必要はない。しかし行動手順が最初から脳にプログラムされているわけではなく、たまたま持っていた習性が、この習性を持たない種よりははるかに高い確率で穴の中の虫をおびき出しゲットする経験につながり、これを自習のように繰り返すことで、一連の動作と得られる結果が記憶に固定化され、道具を使う能力が完成すると考える。
もちろんこの習性があれば、おなじ道具を使う方法を他の個体を見て学習してもいい。この場合はトライアンドエラーを繰り返す自習よりははるかに確実に獲得するだろう。しかし、もし枝を穴に突っ込むという習性がなかったら、この能力は生まれない。
道具を使う能力を考えると、私たちは虫を取るという目的、それを達成するための枝の機能についての表象をカラスが持っていると考える。そして、「普遍文法」と同じで、学習することなくこの能力が獲得されていることを観察すると、全てが遺伝的に決まってしまうと考える。しかし、人間の子供が文法的な言葉を話すのは1年半以上過ぎてからで、それまで様々な経験をする。とすると、アララに特定の遺伝的な習性が存在すれば、あとは試行錯誤を繰り返して道具を使う能力を獲得するように、意味のある統語法を支える普遍文法という能力も、脳の文法特異的な特別回路といったものではなく、様々な領域を統合する能力のような背景に、試行錯誤型の自習が加わった2段階で生まれると考えることも可能だ。このように、獲得に時間がかかる性質は、他の個体や社会からの学習がなくとも、試行錯誤という学習が加わった結果である可能性があることは常に注意する必要がある。
言語と同じで、人間の道具使用の発達についても多くの研究が発表されている。人間も生まれた時には道具を使う能力は全く備えていない。腕や手の運動機能は6ヶ月をすぎると備わってくる。この時から、周りにある様々な物体を触って確かめる試行錯誤により、見つけたものを自分で操作できるかどうかを自習する。この行動は、アララが枝を拾う習性に対応するかもしれない。この見つけたものを手で触って操作性を確かめる行動は一種の遊びに見える。実際、Rat-Fischerらの観察によれば、生後18ヶ月まで道具とおもちゃの区別はない(Rat-Fischer et al, Journal of Experimental Child Psychology 113, 440, 2012)。すなわち、自分の目的と、物体の性質についての表象が機能として表象為直され、それに基づいて行動シナリオを表象できるためには、言語発生と同じだけの時間がかかることがわかる。
言語の発達も、生後半年ほどから訳のわからない赤ちゃん特有の発声が始まり、12ヶ月頃より少しづつ意味のわかる単語の並びが出始め、2年目ぐらいに意味のある文章が出るようになるが、この時間的経過も道具使用の発達経過と似ているように思える。ただ、後に議論するが、言語は社会から離れて完全に自習することはできない。一方、おそらく簡単な道具であれば、社会から離れた人間も道具を使うのではないだろうか。実際、最初の道具が見つかる300万年前にはおそらく言語は存在しなかった。このように、道具の使用と、言語の使用のように、人間に比較的特有の性質は、多くの点で重なると同時に、多くの点で独立している。
このことがわかるもう一つの例が、失語症患者さんの研究だ。後で詳しく説明するが、血管障害などで特定の脳領域の機能が失われた患者さんに見られる言語障害の研究は、言語に必要な脳回路の研究に大きな役割を果たしてきた。これを失語症と呼ぶが、この失語症の患者さんの中に、道具が使えくなってしまったケースが多く見られることが19世紀の終わりから気づかれるようになった。これを「失行症」と呼ぶ (図2)。
このことがわかるもう一つの例が、失語症患者さんの研究だ。後で詳しく説明するが、血管障害などで特定の脳領域の機能が失われた患者さんに見られる言語障害の研究は、言語に必要な脳回路の研究に大きな役割を果たしてきた。これを失語症と呼ぶが、この失語症の患者さんの中に、道具が使えくなってしまったケースが多く見られることが19世紀の終わりから気づかれるようになった。これを「失行症」と呼ぶ (図2)。
図2 失行症の患者さんが櫛を逆さまに向けて使おうとしているところ(出典:Wikipedia)
ほとんどの失行症は、左の後頭頂皮質の血管障害により起こる。患者さんは櫛とは何か、ハサミとは何かが理解できるにもかかわらず、櫛を逆さまにして髪に当てたり(図2)、ハサミを閉じたまま紙を切ろうとする。詳しい解説は避けるが、重要なのは多くの患者さんに、様々なタイプの失語が併発することだ。もちろん、多くの失語患者さんは失行を伴わないし、まれではあるが失語症状の全くない失行症の患者さんも存在する。このように、道具使用と言語能力は、働いている脳領域から見ても、多くの点で重なっていると同時に、多くの点で独立している。
このことは、言語を例えばブローカ領域、あるいはウェルニッケ領域がコントロールしていると局在論的に決めつけることは間違っていることを意味している。道具使用との比較から見ても、言語能力には多くの脳領域が関わり、またそれを統合するメカニズムが存在しているはずだ。
さて次回は、より言語に近い音楽に関する能力を言語能力と比べてみる。
[ 西川 伸一 ]
臨床医学では、一人一人の患者さんから得られる経験を大事する。これは症例報告という形式で発表される論文に現れている。滅多に起こらない病気の場合、ほとんどの医師は直接経験することなく終わる。しかし稀とはいえ、いつなんどき同じような患者さんが診察室にやってくるかもしれない。このためたった1例の経験でも、多くの医師と経験を共有することが重要で、これが症例報告を重視する伝統として生きている。今回は、このような症例報告論文の紹介から始めたいと思う。

図1 引用元、説明などは本文参照
論文のタイトルページを図1に示すが、兵庫県立リハビリテーション中央病院のグループがBrain and Cognitionに昨年発表した論文だ(S. Uetsuki et al. / Brain and Cognition 103 (2016) 23–29 )。53歳右利き女性で、左側頭葉に脳梗塞がおこり、その結果歌うことができなくなった患者さんの報告だ。この症例が報告された理由は、ウェルニッケ領域(前々回参照)を含む左側頭葉に大きな梗塞があるにもかかわらず、失語症はおこらず、音楽の表現だけが障害されている点だ。実際には発作直後の数日、カナと漢字両方の識字障害がみられているが、これはすぐ正常化している。音楽能力が犯されているが(失音楽症と総称しておく)、馴染みの音楽を聞かせると題名を当てることができる、要するに音楽を聞く方の障害はほとんどないが、ピッチの高低の表現、要するにメロディーを歌うことだけが犯されている。
専門家でなくともこの症例から以下のことがわかる。1)脳の局所的障害によって音楽能力が障害されること、2)失語と同じで、音楽能力の障害も、音楽を聞く時の障害と、音楽を表現する時の障害に分かれること、3)メロディーとリズムは全く別の場所によりコントロールされていること、4)音楽も言語もともに人間特有の高次脳機能だが、メロディーを表現する音楽能力と、言葉を話す言語能力とは別々の機能であること、などだ。
前回紹介した失語症や道具使用能力の異常・失行症を思い出していただきたいが、脳イメージングが発達するまで、特定の脳機能とそれを調節する領域の対応関係は、卒中などの脳障害の症例の解析が頼りだった。実際、脳障害の後、あるいは生まれつき音楽の認識や表現する能力が欠損している患者さんの症例報告は19世紀後半から行われている。中でも有名なのは、作曲家ラベルが自動車事故の後、失語、失行、失読、識字障害とともに、作曲する能力を完全に失ったことを記載した症例報告だろう。ラベルの例から、障害によってはこのように、失語、失行、失音楽症が同時に障害されることもある。
実際には、失音楽症の表現は極めて多様で、個人差が大きく、失語症以上に決まった領域にマッピングが難しい。例えばラベルのようなプロの音楽家の失音楽症は左側頭葉の障害による場合が多いことが知られている。一方、多くの失音楽症の症例を集めて検討した研究(例えば2016年、Journal of Neuroscienceに報告された77症例の検討:Sihvonen et al, J.Neurosci. 36:8872, 2016)では、失音楽症の半数に失語が合併しており、言語と音楽能力に関わる共通脳領域の関与を示している。また失音楽症は言語の犯されにくい右側頭葉の梗塞による症例が多いことも報告されており、最初紹介した日本の症例が珍しい例であることを示している。事実、右脳の障害で失音楽症が起こるケースが多いことは、右脳は芸術、左脳は論理という誰もが知っている通説に合致しているようにも思える。これまでの研究は、音楽能力の形成は、言葉と比べてもさらに個人差が大きいことがわかる。最近音楽の様々な要素を経験する時に活動する脳領域を調べる脳イメージング研究が進み、音楽の認識のさらなる複雑性が明らかになってきているが、話が膨大になるので、紹介はやめておく。
これまでの研究をまとめると、私たちは曲を聴くとき、あるいは音楽を表現するとき、一つの統合された全体として認識し、それを表現するが、実際にはメロディー、ハーモニー、リズム、絶対ピッチなどなど、様々な脳領域が別々に働いて集めた情報を、脳内で統合された表象に形成し直して認識している。神経回路的に考えると、それぞれの要素の表象には時間差があっても、正確に統一したリズムの中に統合され、一つの音楽として認識できる過程は、脳を理解するためには格好の課題と言える。しかも、同じような表象を多数の人間が共有できる。幸い音楽を聴かせる課題設定は、脳イメージング技術と相性がいいため、今後大きく発展する分野だと思う。
言語、道具、音楽それぞれの能力を支配する高次脳機能の関わりについて見てきたが、各活動は多くの脳領域が関わり独立に支配されていると同時に、多くの領域で機能的重なりが見られることを理解してもらえたと思う。この機能的重なりは重要で、それぞれの能力が、それを支配する共通の領域を介して相互作用を行えることを意味している。
この人間特有の3つの能力の進化をみると、短時間に急速に多様化、複雑化していることがわかる。例えば、道具は300万年ぐらい前に開発されて以来、5万年前までほとんど変革を遂げることはなかった。しかし私たち現代人の先祖が言語を獲得するや否や、急速な進化が始まった。一方、言葉を持たなかったと考えられる(これについてはいつか議論したい)ネアンデルタール人では、道具の進化をあまり認めることはできない。
一方言語の方も道具により大きく変化する。私たち現代人でも、話された言葉をそのまま覚えておくのは難しい。これを克服しようと文字が誕生するより随分前から、頭に浮かんだ表象を書き留めて覚えるための道具を必要としたはずだ。この方向の道具の進化の中で、言語能力と道具を使う能力の相互作用が始まり、両者は言語を生み出すとともに急速な進化を果たし現代文明を作り上げた。言語と道具は今や、言語翻訳や、言語シミュレーションのような、人間の能力を大きく超える道具を作り出すことにすら成功し、またこの道具により世界の言語が変わろうとしている。

図2 最古の楽器。(出典:Wikipedia)
音楽と道具についても同じことが言える。最初、音楽は体を使って表現されていたはずだ。しかしおよそ35000年前には図2に示すフルートのような複雑な楽器が作られたという証拠が残っており(図2)、道具を使う能力と音楽能力がこの時期には相互作用を始めていたことがわかる。そして今、世界を見渡せば、言語に匹敵するぐらい多様な楽器が存在しているだけでなく、電子化、コンピュータ化などの道具との相互作用により、全く新しい音楽が誕生し、それが脳の成長に大きな影響を与えると考えられている。
この急速な進展は、個人の頭の中で3者の能力が相互作用するだけでは生まれない。次回の話題にしようと考えているが、言語、道具、音楽の3者が共通に持つ重要な特徴が、人間の脳の活動から生まれたにもかかわらず、個人個人の脳活動から独立できている点だ。すなわち、言語、道具、音楽は個体間のコミュニケーションが前提になっており、活動が意味を持つためには脳内に形成される表象が一人の個人の脳内で止まるのではなく、複数(多くの)脳内に同じような表象が形成されていることが必要になる。すなわち、3者に関わる脳活動は、他人(社会)と共有されることで、個人(人間)から独立することが可能になり、人間の活動であるにもかかわらず、人間から独立して進化できる。こうして独立して発展する、言語、道具、音楽を、個人個人は一生を通して学び続ける。このサイクルを繰り返すことで、言語、道具、音楽は進化するとともに、人間の脳回路を変化させてきた。
言語、道具、音楽の各能力が社会に共有されることで個人から独立し、独自の発展が可能になったことが、人類の脳構造はこの5万年ほとんど変わっていないにもかかわらず、3つの能力が急速な進歩をと人間だけが文明を形成できた理由だろう。このことは言語を考える上で最も大事な点で、詳しい議論は次回以降に改めて行う。
今回は最後に、私が音楽と言語を考える時大変参考になった、Steven Mithen著、『The Singing Neanderthal』を紹介する(図3)。

図3 Steven Mithen著『The Singing Neanderthals』のカバー。
もちろん私が紹介するより、音楽と言語の関わりについて興味を持たれる方には、直接読んでもらうのが一番だ。現在絶版になっているが、一度は邦訳(『歌うネアンデルタール : 音楽と言語から見るヒトの進化』熊谷淳子訳)も出版されており、日本語で読むことも可能だと思う。
Mithenの興味は言語の発生だが、「音楽能力なしに言語は誕生しなかった=音楽能力が言語より先に発生した」と考えている。彼が提案する言語発生についての大胆な提案を私なりに改変したものを図4に示す。
図4 Steven Mithenの提案する言語発生に至る過程
チョムスキーと異なり、Mithenにとって、言語はあくまでもコミュニケーション手段から発生したものだ。同じようなコミュニケーションはサルだけでなく様々な動物で見られる(サルの会話の様子はYouTubeに多くの例が掲載されている。例えば、https://www.youtube.com/watch?v=JLOn8F0p96s参照)。そして伝達できるものは感情と一体化した情報だけだ。例えば、乳児と母親とのコミュニケーションを見てみれば、そこで行われている伝達が、専ら感情の伝達に限られているのがわかる。そしてこのために生まれるのが、彼がholistic languageとよぶ、一つのシラブルで伝えたいことの全てを表現する言葉だ。赤ちゃん言葉を例として説明すると、母親の乳房を求めて「mamama」と声を発した赤ちゃんに、お母さんもそれに合わせた赤ちゃん語で答えているとき使っている言葉がholistic languageになる。
感情のコミュニケーションという意味では、音楽も同じだ。言語の混じった歌という形式を取らない限り、ほとんどの音楽家は、音楽が感情を伝える手段である点では一致している。そして、この感情の伝達から、情報の伝達が発展する可能性も、太鼓が楽器としてだけでなく、離れた人間同士の情報伝達に使われるようになることを考えると理解できる。
図4のMithenのシナリオでは、直立歩行を果たし、集団で餌を漁る生活を始めた原始人は、まず交尾のための競争、そして私たちと同じように乳児との対話などに、限られたボキャブラリーのHolistic languageを発展させ、それを仲間同士の情報交換にも使っていた。情報伝達という観点から見ると、Holistic languageは極めて素朴なコミュニケーションにしか使えないのだが、古代原人の生活にとってそれ以上のコミュニケーション手段など実際には必要なかったのだろう。
そして約50万年前、ネアンデルタール人と別れた我々の祖先は、太鼓の音楽を情報伝達に使い始めたのと同じように、holistic languageを分節化し、感情と情報を分離することに成功する。一方、ネアンデルタール人は、ずっとholistic languageを使い続け、行動から感情を分離できないまま、文明の発展が行き詰まることになるというシナリオだ。
ネアンデルタールの遺跡を見ると、埋葬時に花を手向けるなど、豊かな感情表現の痕跡が残っている。しかし感情を伝達する音楽だけでは、文明の発展は限られている。一方、ホモサピエンスではさらに複雑な情報伝達手段が必要になる状況が生まれ、音楽脳に重なる形で、感情と情報を分離し、言語を分節化する能力が生まれる。残念ながらこのきっかけが何だったのかは今も謎のままだ。しかし、一旦独立した言語能力が生まれると、あとはそれぞれの機能が相互作用を繰り返し、現代文明へと突き進む。
このように、言語、道具、音楽という人類特異的能力は個人だけでなく、多数の人間の脳内で相互作用することで独自に進化する。このため、言語の発達を理解する時、この三者を常に念頭に置いて考えることは重要だ。
[ 西川 伸一 ]
これまでの話で、言語の使用や発生が脳科学の問題であることは理解してもらったと思う。しかし前回見たように、記憶、自己、意識についての脳科学と、言語、道具、音楽などの脳科学には、他の個体とのコミュニケーションの必要性という決定的な違いが存在している。今回から、この問題について考えていきたい。議論を進めるにあたって、言語機能を人間特有の機能として考える立場をとり、様々な動物や、「歌う?」ネアンデルタール人も比較しながら議論を進めたいと思っている。
さて、チョムスキーは言語をコミュニケーションの観点から捉えることは、言語の本質を見誤らせると考えていた。これは、彼が言語の最も重要な条件は私達が生まれついて持っている統語能力(普遍文法)で、この人間共通に持つ能力があればおのずとコミュニケーションが可能になると考えたからだ。
たしかに私たちは必ずしも言語をコミュニケーションのためだけに使っているわけではない。ほとんどの人は、物を考える時にも言語に頼っていおり、特に複雑な内容を頭の中で考えようとするとき、言語に頼っていることに気づく。これは、言語が脳内でも覚書として、またチョムスキーが言うように統語に関わる論理的システムとして私達の思考を助けてくれるからだろう。しかしこの考えに立つと、「普遍文法を可能にする神経ネットワークの遺伝的背景」を追求することが言語発生の生物学としては重要な課題になり、基本的には言語の全てが私たちの脳内でとどまっていると唯脳論的に考えることになる。この結果、言語、道具、音楽が持つ全く新しい性質、すなわち私たちの身体的生命から独立できているという性質を見落としてしまう。(実際にはチョムスキーも普遍文法をinternal languageと、それが身体から外化され発展するexternal languageに分けているが、生物学的にこれをどう攻めるかは明らかでない。)
「言語も脳科学の問題と言った後で、言語は身体から独立している、とは何事ぞ」とお叱りを受けそうだ。これまでゲノム誕生に始まって、脳内神経ネットワークまで長々と生命が生み出してきた情報について見てきたが、ゲノム、エピゲノム、シグナル分子、神経ネットワークなどを媒体とする情報は全て私たちの身体とともに消滅していた(現在ではゲノム解読結果をPCに残すことが可能になってきて、この原則は崩れつつある)。一方、言い回しの癖や、発音など個人的な性質を除くと、言語は私たちが死んだあとも、独立した情報として残る(図1)。例えば日本語は私たちの生命を超えて存在し続ける。この意味で、言語は我々の身体から独立している。
最初に述べたように、言語も私たちの先祖の脳内活動から生まれ、また私たちの脳内活動が維持してきたもので、決して天から降ってきた情報システムではない。従って、「言語は脳科学の問題」ということも正しい。ただ、複数の個体間のコミュニケーションのための情報として発達したため、集団(社会)により共有される部分が生まれ、共通部分と個別部分の二重構造になった(図1)。これが個人が消滅しても、言語が続く理由だが、2重構造の成立と維持について、図1を眺めながら日本語を例にもう少し説明してみよう。
図1 言語以外の情報(この図ではゲノム、エピゲノム、神経情報を挙げている)は全て身体に拘束されており、身体とともに消滅する運命にある。言語も最初は脳内の活動の一つとして生まれるが、コミュニケーションのために発達し、複数(社会)で共有できる部分を形成したおかげで、この部分は個人の身体から独立した体系として成立できている。この体系は、常に変化しており新しくなっている。生まれてきた人間は、この新しい体系を習得する必要がある。従って、個々の個人から独立した体系自体も、私たちの脳とは無関係に存在するわけではなく、一生を通して学習され、また共通部分に一部がフィードバックされるという関係を保っている。おそらくこの言語の2重構造が、クオリアといった主観と客観の2元論の背景にある。個人から独立し、社会で共有できる言語の体系は現代文明のルーツで、今やその成果としてのゲノムプロジェクトを介して、本来なら身体に拘束されているゲノムやエピゲノムを身体から離して保存することに成功している。
私の頭の中には日本語の全てが存在しているわけではなく、ほんの一部が存在しているだけだ。すなわち、私が死んでも日本語はビクともしない。この意味で、日本語は私の身体とは別のところに存在している。そして私は生後すぐから現在まで、この日本語を学び続けることで、私の中の日本語を形成・維持・変化させいる。このように、日本語は個人から独立して存在していても、常に人間の脳と相互作用を続けている。言語とは新しく生まれた個人がそれを習い、使い続ける中で初めて維持されるシステムだ。将来もし誰も日本語を使わなくなれば、日本語という記録は残るだろうが、その時点で日本語の変化は停止する。図1はこの言語の2重構造(一人一人の個人の頭の上に小さな言語を置いた上に、共有部分としての大きな言語を描いている)を示している。
色の認識を例にさらに考えてみよう。20世紀米国の分析哲学ではクオリア問題がよく議論された。これは私が赤色を見た時の主観的体験を客観的に、すなわち他の人に理解できる形で定義できるかについての議論だ。私は、この議論は図1に示した言語の2重構造を反映していると思う。考えてみると、私たちはそれぞれ赤色について主観的質感を持ち、個別に定義する。ただ、私のクオリアは、言語を学習する過程で言語の共通部分にある「赤色」という言葉が対応して初めて成立する。一方、言語の共通部分も多くの個人の赤色の体験からフィードバックを受けて変化し続けている。このような言語の2重構造のおかげで、実際には赤色に対する個人の質感と、言語の共通部分が代表する赤色との差など何ら意識することなく、「赤色」について多くの人と客観的に議論できる。
この言語の2重構造は、言語が独立して進化する原動力でもある。私たち個人はこれまで誰も経験したことのない新しい経験を日々繰り返している。この新しい体験は、当然言語化され、言語の共通部分へとフィードバックできるが、実際にはほとんどがフィルターされ、個人の経験で終わる(これは私たちの感覚が日々膨大なインプットを経験しながら、フィルターされ選ばれたほんの一部が神経ネットワークの自己を書き換えるのと似ている:76話)。
次に、例えば空飛ぶ円盤を見た体験がフィルターを通って共通部分へとフィードバックされる条件を考えてみよう。これまで飛ぶ円盤を見た人は誰もおらず、あなたしか見た人はいないとしよう。当然空飛ぶ円盤を表現する言葉は言語の共通部分にはまだ存在しない。この時、「空飛ぶ円盤」が言語の共通部分を書き換えるためには、この体験をまず言葉で表現する必要がある。この過程で「空飛ぶ円盤」という最適の言葉に思い至り、めでたく共通部分に新しいボキャブラリーが誕生する。
この例からわかるのは、私たちの言語が、あらゆる概念を現存の単語を使って表現できるとてつもない力がある点だ。このことは、広辞苑でも大辞林でも、皆さんが使っている辞書を思い浮かべると理解できる。そこには日本語として使われるほとんどの単語が、他の単語で表現されている。これが空飛ぶ円盤を見たあなたが、「空飛ぶ円盤」という新しい表現に到達でき、新しい概念を「空飛ぶ円盤」という言葉として言語の共通部分にインプットできる理由だ。言語を構成する各要素は、脳内神経ネットワークと同じように、意味のネットワークを形成している。このネットワークが、個別の脳につながる言語システムからインプットうけ、不断に書き換えられている。
この2重構造に基づく進化力のお陰で、毎日の生活での言語体験がどんなに乏しく、またそっけなくとも、言語はますます豊かになることができる。このことは会話を主体とする戯曲と、小説の文章を比べるとすぐわかる。
少し古いが菊池寛の「父帰る」の一場面の会話を抜き出してみよう。
新二郎
ただいま。
母
やあおかえり。
賢一郎
大変遅かったじゃないか。
新二郎
今日は調べものがたくさんあって。
ああ肩が凝った。
母
さっきから御飯にしようと思って待っとったんや。
賢一郎
御飯がすんだら風呂へ行って来るとええ。
新二郎
たねは。
母
仕立物を持って行っとんや。
なんとそっけないことか。複雑な構文もないわけではないが、これももっと単純な構文に書き直せるだろう。それで用は足る。
実際、日常会話で長々と文章を述べようとしても、すらすらと文章を述べられる人はまれだ。私自身、対談の原稿が上がってくると、なんと意味不明の文章や、「あの」「あれ」「それ」と言った単語にならない単語を連発しているのに気付き、いつも幻滅する。結局、私たちの日常生活はこのようなそっけない乏しい言語の使用で終わっている。しかし、小説や、戯曲でも長いセリフを読むとき、私達の言語がいかに複雑で、高いレベルに達しているかよくわかる。これは個人の言語体験を超えて、言語の共通部分が急速に進化していることを物語っている。
説明が少し長くなったが、この言語の2重構造が、言語の最も重要な特徴で、この構造は言語がコミュニケーションを目的としているからこそ可能になった。この意味で言語の原点はコミュニケーションだと私は思っている。
ではこの構造はどのようにして生まれたのか?この言語誕生の問題に関しては、前回紹介した「歌うネアンデルタール人」の著者Steven Mithenの考え方に私も強く影響されており、彼の説を下敷きに考えていきたいと思っている(これ以後、言語という言葉を、言語発生前のプロト段階と発生後の言語の両方を含む意味で使うので注意してほしい)。
彼は言語の発生を考える時、コミュニケーションのために使われる体系をmanipulative languageとreferential languageの2段階に分けている(図2)。

図2:Mithenの考え方に従った言語発生の2段階。
いずれもコミュニケーションのために発達し、最初は一つの音の塊に、他の個体の行動を促すすべての情報が備わったholistic & manipulative languageだが、その後フレーズが単語に分節され、各単語が特定の概念を参照する、segmental & referential languageへと変化する。segmental languageはcompositional language(合成的言語)と読み替えることもできる。
Manipulative languageのわかりやすい例としては、赤ちゃんの発声を考えればいい。泣き声のことも、「ママママ」などいろんな発声があるだろうが、それを聞いた大人は何を望んでいるのか発声の意味を解釈し、何とか希望を叶えようと努力する。もちろんベテランのお母さんになると、声を聞くだけですぐに解釈することができるが、いずれにせよその発声により周りの人間が操作を受けることになる。これがmanipulative languageだ。
Manipulativeな発声は人間特有のものではなく、もともと多くの動物や鳥類で見ることができる。ヒョウ、ワシと危険のもとを区別して警告を発することで有名なベルベットモンキーの鳴き声を効いた仲間が安全なところに逃げるのは(YouTube参照:https://www.youtube.com/watch?v=hEzT-85gEdA)、それを聞いた仲間を適切な場所に逃がすためのmanipulative callの典型例だ。この鳴き声は一見ワシ、ヒョウと特定の危険の原因を参照(referential)しているように見えるが、声が単語として特定の動物を表象しているかどうかはかなり疑問だ。実際には、「空から危険が迫っているので地下に潜れ」のような、一言で全てを命令するholisticな内容を持つ鳴き声と言える。同じように、赤ちゃんの発声も「お腹が減った、おっぱいが欲しい」のようにひとつのフレーズが分節されないで表現されるholisticな言語だ。
Holistic & manipulative languageの構造がピッチやリズムの変化だけで表現される音の塊の上に、他人の行動を促す強い感情がかぶさっている点で、音楽と多くの共通性を有している。そのためMithenは、この段階のプロト言語が音楽と一体となって進化したと考えている。この考えに基づいて、彼は前回の図4で紹介したように、直立原人からネアンデルタール人までの進化過程で、Holistic languageは音楽とともに十分な進化を遂げ(複雑化、多様化)、例えば獲物のモノマネのなどのジェスチャーを組み合わせれば、ネアンデルタール人がグループで大型動物を狩る時のコミュニケーションを十分まかなうことができたと考えている。
もちろんこれが正しいかどうかは見てきたわけではないし、また遺物が残っているわけではないのでわからない。しかし、例えばレンジャー部隊が手の合図だけで、複雑なタスクを実行してしまうような映画を見ると、Hoistic languageの力は十分位理解できるし、十分あり得る話だと思う。
しかしこうして生まれた manipulative languageによるプロト言語段階はホモサピエンスの誕生とともにreferential languageへと変化して、今回議論したような、身体から独立した進化する言語体系が可能になった。強く感情に支配されるHolistic languageは身体から独立できず、進化の速度は極端に遅い。次回から言語発生にとって最も重要な段階に進んでいくが、この問題を考える前のコミュニケーションの基本条件として理解しなければならないのが、「他人も自分と同じように考えることができることの認識」、すなわち「Theory of Mind」の問題だ。次回はこれについて考えることにする。
[ 西川 伸一 ]
今回から2回にわたって言語発達と社会性について考える。
何歳ぐらいから私たちは周りの人の顔色を伺って過ごすようになるのだろうか?今現役を退いて、部屋でゆっくり論文を読んだり、原稿を書いたりして、あまり人とは接しない時間を持てるようになると、自分が人の顔色を常に伺っていることを実感する。一人の時間が終わると、家族や友人は言うに及ばず、街中で多くの見知らぬ人と出会う。行きすがりの人について一々深い空想を巡らせるほど偏執狂ではないが、顔をチラッと見て「急に襲ってはこないか」など無意識に判断している。ましてや満員電車に乗ろうものなら大変だ。周りの人をそれとなく窺い、争いが起こらないように、あるいは痴漢と間違えられないよう、最適のポジションを探してしまう。
家族ならある程度心が読めるが、赤の他人の心の中などほとんど何もわからないことがわかっているのに、人間は他人が何を考えているのかを知ろうと、多くの努力を払っている。程度の差はあれ、このように日々他人の心を読もうと常に努力するのは私だけではないはずだ。というのも今の世の中では、このような意図が欠けている人は、病気だと診断される。例えば自閉症スペクトラムと診断された人たちがその典型だ。
現在では、自閉症スペクトラムは病気ではなく脳の多様性として捉えることが普通になっている。この観点に立つと、自閉症スペクトラムの人は、他人の心を読もうとする悲しい人間の習性から解放された人たちということができる。ただ、問題は自閉症の人たちの多くが、言語障害など様々な社会への適応障害を同時に併発している点だ。従って、脳の多様性だと放っておくことはできず、社会と折り合いをつけるためのプログラムが用意される。プログラムの多くは、できるだけ早期に診断して、脳回路がフレキシブルな発達期のうちに介入して言語や社会性を回復させるよう計画されている。そこで、フロイトをもう一度思い出しながら、乳児の脳の発達を見てみよう。
脳科学的な表現を用いて言い換えると、フロイトは自我を、生まれたばかりの脳が持っている自己基盤が、体験により書き換えられた結果と考え、特に幼児期の体験の重要性を強調した(第78話)。もちろん脳の自己の書き換えは一生を通じて行われており、自我の形成へ影響を持つ体験は幼児期に限らない(PTSD:心的外傷性ストレス障害はその典型だ)。ただ、幼児期にインプットされる体験は、外界からのインプットをほとんど受けたことのない脳に対する最初の体験である点、またインプットの種類が限られている(と私たちは思っており)点で、成長後の体験と比べるとそのインパクトは相対的に大きい。逆に言うと、体験を重ね、書き換えが進むと、体験のインパクトは低下するため、一般的な体験の自我形成への影響は低下する。繰り返すが、感覚器の発達していない乳児の体験は、個体間での違いが少ない。事実生後2−4ヶ月まで、耳は聞こえても成人の感覚入力の8割を占める視力はほとんどないと考えていい。当然母親の顔をはっきり認識はできない。このため、最初の経験は、音、匂い、そして母親との直接コンタクト、それも乳房に対する皮膚感覚や味を介して入ってくる。インパクトの大きい最初の体験は、個体間の多様性が限られているからこそ、人間の自我が持つ共通性の重要な要因になっている。フロイト的に言うと、「自我形成には、人類共通に「口唇期」の母親の体験が重要である」となる。
フロイトはこの体験が決して受身的なものでないことを見抜いており、これを「対象備給」と呼んだ(第78回)。すなわち、乳児が目の前の対象に注意を払うことが、欲動にドライブされる能動的な過程であることをこの特殊な言葉で表現した。この力動を欲動というより本能といった方がいいかもしれない。乳児期にだけ「口唇追いかけ反射」とよばれる母親の乳房を探す反射が存在するが、これも能動的に体験を求める一つの駆動力になっている。実際この外界への能動性が乳児の特徴と考えられている。まだ視力が完全でなく物の区別がつかない3−4ヶ月までに、正常の赤ちゃんは必ず相手の顔をじっと見つめるようになるし、動く物体を追いかける。また、音がする方向に顔を向ける。このように、視覚が発達する時期までに、能動的に体験を積み重ね、最終的には家族とそれ以外を見分けることができるようになっている。フロイト的には、この過程こそが母親、父親と、自分の関係を理解し整理し、母親への欲動の抑制などが始まる重要な時期になるが、今回は議論せず先に進む。
図1は、米国疾病予防管理センターが、乳児の発達異常を発見するために公表しているチェックリストだが、もし乳児が音に反応せず、動くものを見つめず、笑わず、口に指を持って行かないならすぐに医師に相談せよと強い口調で警告している。もちろん自閉症スペクトラムが診断の対象ではないが、能動的に体験を求める行動が見られないことは、自閉症スペクトラムを含む、発達障害が強く疑われることを示している。

図1:米国疾病予防管理センターから出ている、発達障害早期発見のためのチェックリスト。
診療現場では、社会性の欠損、反復行動、そして言語の発達障害が自閉症スペクトラムの最も重要な診断基準になっている。図2は、Autism Speaksと呼ばれる自閉症スペクトラムの患者さんのために活動している団体のサイトから転載した表で、自閉症スペクトラムでみられる主要な症状、および関連する疾患が描かれている。
図2:自閉症最大の患者さん支援団体Autism Speaksが示している自閉症の主要症状と、他の様々な異常との関係を描いた図。
出典:http://www.autismspeaks.ca/about-autism/signs-and-symptoms/
この表から自閉症スペクトラムがいかに一筋縄でいは扱えない、多様な状態の集まりであるかがわかるだろう。特定の遺伝子変異に起因する一部の症例を除くと、自閉症の表現形はほとんど個性と言ってもいいほど多様だ。事実、最近急速に進展しているゲノム研究では、自閉症スペクトラムと関連する遺伝子はすでに100を超え、特定の遺伝子だけで説明することは難しい。しかし、早期発見のための乳児の症状チェックリストをよく読むと、自閉症の背景には能動的に新たな体験を求める衝動の低下があるように私には思える。すなわち、他人の顔を伺い、心を読もうとする衝動を形成する脳回路が最初から低下している。事実、多くの研究は、自閉症スペクトラムの人は脳回路の構築が異なっていることを示唆している。例えば、扁桃体に留置した記録電極を用いた研究では、普通多くの神経が目に反応するのに反し、自閉症では口に反応することが明らかにされている。すなわち、自閉症では神経細胞レベルで刺激反応性が異なっていることを示している。
ただ、脳の状態が症状として現れるのは、これらの単一神経レベルの反応が統合された結果で、これを脳科学の言葉で説明するのはまだ簡単でない。例えば、先に挙げた自閉症児で低下している他人の心を読もうとする衝動と能力は、人間特有の性質として研究されているが、その神経科学的本態の理解にはまだまだ研究が必要だ。この能力をmind readingと分かりやすい形で表現することもあるが、心理学や哲学ではこれをTheory of Mind(ToM)と呼び、様々なテストが開発されてきた。中でも自閉症を理解するためのテストとして研究に使われるのが、False-beliefテスト(他人の間違った考えをわかるテスト)だ。
ToMテストの中で最も有名なのが「サリーとアンのテスト」とも呼ばれているテストだ。図3に示すように、このテストでは、2人の女の子が登場する一連の場面を理解させる。
「同じ部屋に、サリーはバスケットを、アンは箱を自分の持ち物をしまう場所として持っている。ある時サリーはアンの眼の前で自分のボールを自分のバスケットにしまう。サリーが去った後、アンはそのボールを自分のボックスに移して部屋を去る。その後サリーがボールで遊ぼうと部屋に帰ってくる。」
この状況を理解させた後、
図3:ToMの能力を知るための定番都なっているサリーとアンのテスト。フランス語版Wikipediaの図を改変して示している。
質問に対し、3歳児までは実際に自分で見たことだけが優先され「ボールは箱にある」ことはわかるが、サリーがどう考えるかは理解できていないことが普通だ。しかし4歳児になると、「ボールは実際にはアンの箱の中にあるが、サリーはバスケットの中にあると思っている」ことがわかるようになる。ところが自閉症スペクトラムの子供は、4歳児をすぎても今ボールが箱の中にあることはわかっても、サリーがどう考えているかを理解できない。すなわち、他人の立場で考えようとするモチベーションがない。自閉症スペクトラムの症状は多様で、言語能力も個人により違っているが、自閉症スペクトラムの人に限って調べた研究でも、ToMと言語能力が相関することも明らかになっている。
余談になるが、最近このサリーとアンテストを、類人猿で行った論文がScienceに掲載された(Krupenye et al, Science, 354:110)。詳しくは紹介しないが、京都大学のチンパンジー施設やドイツマックスプランク研究所で行われたビデオを見ると、サリーとアンのテストがそのままサルでも理解できる様改造され利用されているのがわかる (https://www.youtube.com/watch?v=1s0dO_h7q7Q)。
さて、自閉症スペクトラムの児童研究からToMの発達と言語能力の発達が強く相関していることをわかってもらったと思うが、ではなぜこの様な強い相関がToMと言語間に生まれるのだろう。 ToMは、言葉が話せる様になった後に確認できるようになるため、自閉症児でのToMの障害は、言語能力の障害による二次的な結果ではないかという考えがある。もちろん否定はできないが、口唇期に始まる乳児脳の発達過程で体験を求めて外界へ注意を向ける本能や欲動が、自我の形成に大きく関わっていることを考えると、両方の障害はこの発達期に経験を求めて外界を注視する本能・欲動の低下に大きく影響されているのではないかと思える。図2に示した自閉症スペクトラムのもう一つの重要な症状、繰り返し行動も、外界への能動的注視が抑制され続けた結果、変化を嫌うようにったと考えることができる。
個人的な仮説に過ぎないが、もしこの本能・欲動の回路発生段階での形成が自閉症スペクトラムの子供でうまく形成されないとしたら、他人の気持ちを理解するという動機や、言葉を話そうとする動機も失われるだろうし、そもそも変化自体を嫌う性質が前面に出て、繰り返し行動が目立つようになる。こう考えると、自閉症の重要3症状を納得することができないだろうか。 以前人間の行動の動機を支配するのは、結局感情であることについて説明した。実際、自閉症スペクトラムの成人の多くがアレキシサイミア(失感情症)を併発する確率が高いことが知られている。さらに機能的MRIを用いる研究から、自閉症スペクトラムでは感情のサーキットの一端を担う島皮質の活動が低下していることも示されている。このように能動的に体験を求める本能・欲動の障害を、感情障害の一つの表れと捉えることは可能だ。
しかし一方で、自閉症スペクトラムの人たちは音楽を通した感情の伝達や受容については普通の人と同じかそれ以上であることが多くの研究により明らかにされている。さらに、言葉の障害があっても、歌を通してであれば言葉を話せる例があることも報告されている。音楽も感情の伝達であることを考えると、音楽を介する感情の伝達は、自閉症スペクトラムの感情の表現や理解の障害に関わる島皮質とは異なる領域に支配されていると考えられる。いずれにせよ、音楽を介する感情伝達がほぼ正常であるということは、自閉症スペクトラムでは感情が欠如していると考えるより、感情の表現や受容のための一つの回路の形成がうまくいっていないと考える方が良さそうだ。現在では、この事実を利用して、音楽を介する感情の伝達回路を使って、自閉症スペクトラムの子供と交流し、できればもう一つの回路も回復させられないか、いわゆる音楽療法が盛んに行われている。
少し長くなりすぎたので、今回はここで終わる。次回も言語と社会性の問題について考えるが、まず今回残してしまった、自閉症スペクトラムの逆の症状を示す発達障害、すなわち人懐っこい社交性を持ち、知能に比して高い言語能力を持つWilliams症候群から始めたい。
[ 西川 伸一 ]
前回から続いて、言語と社会性について考える。
前回、言語と社会性の発達が遅れ、反復行動を示す自閉症スペクトラム(ASD)を例に、他人の心を理解する能力theory of mind(ToM)に代表される社会性が言語能力に大きく影響し、またこの能力が生まれた時から脳が本能的に持っている能動的に外界へと経験を求める力動を条件にしている可能性について考察した。外界へと積極的に新しい経験を求める本能や情動は(モティベーションと言っていいかもしれない)、フロイトが自我形成に関わる力動として定義した「対象備給」に相当する。フロイトが考えたように、自我は最初から備わっているわけではない。誕生時に備わった最初の自我基盤を、能動的に得られる新しい経験によって書き換えることで形成される。従って、得られる経験の違いは自我の傾向に大きく影響するし、自我形成過程で経験を求める回路の一部が閉ざされてしまうと、形成される自我の安定性や構造は大きく変化する。
以前述べたように(意識と無意識*http://www.brh.co.jp/communication/shinka/2017/post_000007.html)、すべての経験を自我の書き換えに使えるわけではない。毎日体験する膨大な神経インプットの中からほんの一部が選択され(意識過程)、それが自我の書き換えに使われる。この時どの経験を自我の書き換えに利用するかを選択するフィルターの一部は、書き換えられつつある自我そのものである点が重要で、この結果、初期に形成された自我の持つ傾向は、成長とともに増幅されていく。
ASDは外界へ向く力動の低下で、外界とのコミュニケーションに関わる多くのチャンネルが細くなっていると考えられるが、前回述べたように音楽を介する感情の表現や理解は正常以上に保たれている。このことは、自我形成に必要な外界とのコミュニケーションのためのチャンネルは多様で、太さも個人差が大きい。それぞれのチャンネルの総和が幼児期の個性を作る重要な要素として働くが、このチャンネルの種類や太さが遺伝的な原因で特定の偏りを示す場合、形成された自我に共通の特徴が現れることになる。
その一つの典型が、一見自閉症とは全く反対の症状を示す病気ウイリアムズ症候群(WS)に見られる。この病気は1961年にニュージーランドのWilliams医師らにより、大動脈弁の上部の狭窄と特徴的な行動異常と知能障害を示す発生異常としてCirculation誌に発表された(図1A)。
図1 A) Williams症候群が初めて疾患として定義されたCirculation の論文。 B) Creative Commonsとして登録されているNikitina等がActa Naturaeに掲載した総説論文。写真は全てこの論文から転載している。右端には、WSの子供による手本になる絵の模写を示している。一つ一つのアイテムは認識できても、その空間的位置関係の理解が失われているのがわかる。(Nikitina et al, Acta Naturae, 6,9, 2014)
無味乾燥な医学的表現で症状を列挙すると、1)時に致死的な大動脈の狭窄、2)特徴的な容貌、3)高カルシウム血症、などを中心とする身体的な異常と、精神的異常として、1)空間的バランスの認識異常(図1:簡単な絵もうまくコピーできない)、2)IQ50程度の知能障害、3)高い社会性、4)怒りを理解する力の低下、4)正常以上のボキャブラリーや多弁、などが主症状として挙げられる。身体症状はもっと多彩で、例えば糖尿病の発症も多くのケースで見られる。後で述べるが、この多彩な症状はWSが7番染色体の大きな欠損により発症するためだ。ただ、ここでは精神症状に絞って見ていく。
残念ながら私も症例を見た経験がないので、生き生きと表現するのは難しいが、WSの行動についてもう少しわかりやすく説明してみよう。図1Bはロシアの研究者の総説から転載したWSの子供たちの写真だが、これを見ると,年齢にかかわらず皆楽しそうに笑っている。この人懐っこさは外見だけではない。「誰にも愛され、誰もを愛し、チャーミングな性格」と表現されるように、見知らぬ相手でもじっと目を見つめ、近くに寄って話しかける。同じ総説によると「In WS, the gregarious personality is characterized by a consistent increased interest and approach to strangers, …」と表現されており、「見知らぬ人(もの)に対して興味を持ち続け、近づこうとする」性質を持っている。これは外界からの経験を求める力動が低下したASDのまるで逆で、WSの子供は一般の子供以上に、外部と関わろうとする強い力動が発達していることがわかる。言い換えると、外部へのチャンネルが太いと言える。
この積極性はかなり早い時期から見られることが報告されているが、WSの子供が相手の怒りを理解できないことは、WSを理解するヒントになる。ASDの場合最初から人を避けるので問題はないが、WSの人懐っこい性質は無防備と裏腹で、危険を伴う。このことから、経験を求めて外へ向く本能は、ある程度抑制されており、ASDの場合はそれが強すぎるが、逆にWSではその抑制がはずれれており、人懐っこい性格につながっているのかもしれない。正しいかどうかはわからないが、わかりやすく理解するために、ASDの子供で細っているチャンネルの少なくとも一部がWSでは太くなっていると勝手に考えることにしている。
しかしこの考え方がまんざら間違っていないことが、脳イメージングによる研究で示されている。私たちの行動のモチベーションに関わる脳回路として、前頭前皮質と辺縁系の扁桃体、腹側線条体を結ぶ、褒美のシグナルを発生させ満足させる回路が存在し、感情と感覚や行動を支配している。なかでも扁桃体は表情から感情を理解する過程に関わる領域で、例えば見つめ合う行動にこの領域は必要だ。ただ、小さな扁桃体にも13の核が存在し、ASDやWSの行動を扁桃体全体の活動状況と照らし合わせることは難しい。それでも、ASDでは扁桃体は大きくなって、興奮しやすいことが知られている。一方、怒りの症状に対して普通なら反応する扁桃体が、WSではほとんど活性化されない。想像をたくましくすれば、扁桃体の一部の活動が、外への動機づけのチャンネルの太さの調節に関わっており、ASDでは活性が高いため抑制され、一方WSでは活性が低いため抑制が効かないと考えることもできる。
ただ、これは外に向いたチャンネルのほんの一部だ。例えば、WSの子供は音楽能力が優れていることが知られており、この点ではASDと同じだ。このように、外部に向いたチャンネルの中には、音楽を介する感情のチャンネルのように、WS、ASD共通に太くなっているものもある。一方、前回議論したToMはASDだけでなく、これほど人懐っこいWSでも形成が遅れている。このことから、外部に向けられたチャンネルがToMの発達を決めるのではないこともわかる。
社会性の生物学の対象としてWSに注目が集まっているのは、7番目の染色体上の、7q11.23に構造的に非対立性組み替えが起こりやすい領域があり、ほぼ全てのWS症例でこの部分の欠損が見られるからで、欠損遺伝子の機能解析から社会性を分子生物学的に明らかにできるのではと期待されてきた。WSではほぼ例外なくこの領域の欠損があるが、WSには遺伝性はなく、両親は遺伝的にも正常だ。生殖細胞の発生過程や極めて初期の胚に新たに欠損が起こった結果症状が生まれると考えられている。
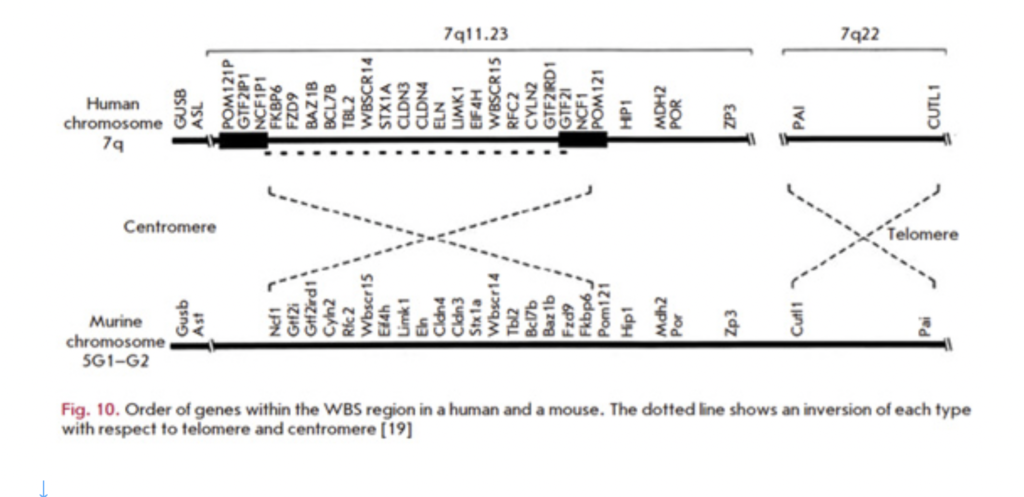
図2:7q11.23上にあるWSで欠損する領域(破線で欠損部分を示している)と、対応するマウスゲノム領域。NIkitinaの総説から転載。
WSのほぼ全てのケースで同じ欠損が見られとしても、実際には人懐っこい性格などの社会性を遺伝子から説明するのは簡単ではない。というのもWSでは欠損は片方の染色体だけに限局しており、さらに、欠損場所には26-28種類の遺伝子がコードされている(図2)。すなわち症状を理解するためには、遺伝子発現量の違いで起こる細胞レベルの変化をとらえ、それを脳の複雑な機能と対応づけるという難しい課題を克服する必要がある。
WSの遺伝異常を症状と関連づける目的で現在行われている研究の方向性の一つは、各遺伝子のノックアウトマウスの作成で、図3に示す様に、WSと関連がありそうな形質を示すマウスが作成されている。

図3 WSで欠損する遺伝子のマウスでの機能。
図3に示した様に、FZD9、STX1A、LIMK1,CLIP2、GTF2iなどが欠損すると、確かに様々な脳高次機能の異常が認められる。特にLIMK1, CLIP2では明確な空間認識の異常が検出されており、WSの症状のモデルになる可能性がある。また、FDZ9ノックアウトマウスでも海馬の細胞死の亢進、癲癇発作の閾値の低下とともに、やはり空間認識障害が報告されている。しかしほとんどの研究は遺伝子が完全に欠損したマウスの解析で、WSの様に片方の染色体だけで遺伝子が欠損するhemizygousマウスの解析はほとんど行われていない。おそらく、明確な異常が検出できないと考えられる。しかし、hemizygousでも変異が組み合わさった場合は、十分異常が検出できる可能性は大きく、今後の研究が待たれる。一方完全欠損が胎生致死のGTF2iについては、hemizygousマウスの行動解析が行われており、WSと同じで、初めて出会ったマウスに対しても警戒心が低下していることが報告されている。
以上の様にノックアウトマウスの解析からWSの症状理解のためのヒントが得られることは間違いないが、WSの多彩な症状をマウスの解析結果のみから推察することは難しい。そこでもう一つの方向として、WSで欠損する大きな領域の中の一部だけが欠損したWS症例を探す努力が並行して行われている。例えば、図2で示した領域のうち、STX1を残してそれより右側の遺伝子だけが欠損している場合でも、完全なWS症状を示すことが報告されており、WSにはSTX1より左側の遺伝子はあまり重要な役割を持っていないことが示された。また、LIMKは欠損していてもGTF2iが欠損しないケースでは、心臓の奇形と空間認識異常は見られるが、精神発達は正常であることが報告されている。この様に、不十分とはいえ少しづつ遺伝子と症状との対応が明らかになってきており、これまでの研究を総合すると、GTF2iに近い領域がWSの精神症状に深く関わると言っていいだろう。
そんな矢先、驚くべき論文がプリンストン大学からScience Advances7月号に発表された。この研究では、オオカミから犬へと家畜化が進む過程で獲得される人間に対する高い社会性をもたらせたゲノム変化の特定が試みられた。そして驚くことに、WSで欠損するGTF2iとその隣のGTFIRD1のコーディング領域の変異による分子の構造変化がこの社交性を説明することが示された。

図4:犬とオオカミを比べて、犬の社交性に関わる遺伝子がWSで欠損している遺伝子であることを示した論文(vonHolt et al, Science Advances e1700398, 2017)。
この研究も合わせて考えると、社交性に関わる脳回路形成にGTF2iとその近くの領域はますます重要になったと結論できる。
最後にWSの言語発達についてまとめておく。発表当初から、WSの多弁を含む高い言語能力に言語学者は大きな興味を抱いてきた。この結果、言語発達や起源についての著作には必ずWSの言語能力が記載される様になっている。ただ、研究が進むとWSには多くの精神発達障害が重なっており、言語能力についてもこれらの障害の影響が大きく、単純な枠組みで理解することは難しいことがわかってきた。このことを認識した上で、あえて単純化してWSの言語能力をまとめてみると次の様になるだろう。
問題の重要性から、言語と社会性についての話が少し長くなってしまったが、最終回は、「社会で共有する言語にアクセスできずに育った個人はどうなるのか?」という問題を考えよう。
プサメティコス王の禁じられた実験
「言語は習わなくとも人間の脳機能として本来備わっている」という命題を確かめる方法が一つある。生まれたばかりの何人かの子供を、社会から隔離し他人との言語によるコミュニケーションが全くない状態で育ててみて、子供達だけが何らかの言語をコミュニケーションに使うようになるか調べることだ。どんな形であれ、もし彼らが共通のコミュニケーション手段を開発できれば、言語は習わなくても人間さえ集まれば自然に発生すると結論できる。しかしこの実験は理論的に可能でも、もちろん禁じられた実験だ。
ところがギリシャの歴史家ヘロドトスは、そんな禁じられた実験をした王様の話を書いている。それがエジプト第26王朝のファラオ(王)プサメティコス1世(紀元前664-610年)で、プサメティコスの禁じられた実験として知られている。プサメティコス1世は地球上に最初に誕生した言語は何か知る目的で、生まれたばかりの双子を誰も住まない島で言葉を全く使わずに成長させ、最初に何語を話すかじっと待った。そして2年目、ついに子供達が最初に発した単語が「bekos(フリギア語でパン)」だったので、最初の言語はフリギア語だと結論したという話だ。この話は、人間は習わなくとも言語を話すことができると古くから考えられていたことを示すエピソードだが、歴史家ヘロドトスの記述とはいえ、本当にあった話かどうかはわからない。
図1ミルクを作るプサメティコス1世のレリーフ(出典:Wikipedia)
この話を言語の二重構造性の観点から考えると、社会で共有する言語に全くアクセスできなくとも私たちの脳の延長として存在している言語を介して他人とコミュニケーションを図る本能により、社会で共有できる新しい言語を自然発生させ得ることを意味する。従って、プサメティコスの実験を「社会で共有される言語に全くアクセスできずに発達すること」と読み替えると、わざわざ人間を隔離する禁じられた実験は必要なく、世界を見渡せば同じような実験は現実に進んでいる。その一つは、幼児期に社会から完全に隔離された子供(野生児:Feral child)の言語発達で、もう一つは生まれつき完全に聴力が失われ、既存の言語から隔離されたろうあ者たちの言語だ.
野生児

図2インドで発見された野生児Kamala。最終的には2本足で立ち、普通の食事をするようになったが、単語は50語を覚えるのがやっとだったとされている。(出典:Wikipedia)
古くから動物に育てられた野生児の話は数多く知られている。もっとも有名なのは1920年インドで発見された狼に育てられたと称するKamalaとAmalaで(図2)及び図3右端の本に詳しく記載されている。発見後9年たってようやく50語を話すようになったことが記載され、社会から隔離され言語が全く存在しない条件で育てられると、人間社会に戻っても言葉を学習するのが難しいことを示している。
さらに最近では1972年にインドで発見された、やはり狼に育てられたと称するShemdeoについての研究が報告されており、1985年に亡くなるまで、一言も人間の言葉を話さなかったことが記載されている。
生後間も無く社会から隔離された(?)ケースをまとめると、最初は4足歩行で生肉を食べ、もちろん言葉を話すことはなく、人間に対しても強い警戒心を持つ。ただ時間が経ち、人間への警戒心がとけると、2足歩行や食事などは一般人と同じ様に振る舞えるまで回復するが、言語だけはほとんど回復しないことが共通だ。これらの話は一般の関心も高く、図3に例示するように、ノンフィクションとして繰り返し出版されている。

図3:動物に育てられた子供についての本は数多く出版されている。他にも、親のネグレクトで社会から隔離された育ったGenieについてのノンフィクションも読むことができる。(出典:Amazon.co.jp)
ただこれらの例は本当に狼によって育てられたのか、発見されるまでの状況について確たる証拠がないことや、科学的、心理学的な研究がしっかり行われていないことが多く、そのまま結論を鵜呑みにするのは危険だと思う。
これに対し、幼児期から極端なネグレクトをうけて社会から隔離され育ち、13歳で発見された子供Genie(図3右写真)については詳しい科学的調査が行われ、研究論文も多く発表されている。Genieの場合は、13歳まで全く言語から隔離されていたにもかかわらず、発見後の訓練で言語能力はおどろくべき発達をとげたことが記載されている。
野生児の観察から、人間の言葉の学習には重要な時期があり、その時期をすぎると言語を覚えることは人間でも難しいと結論された。事実、鳥がさえずる能力を獲得するためには決まった時期に鳴き声を学習する必要があることは、古くからよく知られた事実だ。しかし、Genieの例は、少なくとも人間に関しては、言語習得に絶対的な時期的制限はないことを示唆している。ただGenieの例からだけで、幼児期の学習が言語習得に必要ないと結論していいかは疑問だ。すなわちGenieが既存の言語からどの程度隔離されていたかについては、関係者の証言から想像する以外に方法がないことだ。いくら周りから謝絶された小部屋に隔離されていたとはいえ、部屋の外では様々な言葉が聞こえたはずだ。また、動物に育てられた子供と違い、ネグレクトの場合は、無言のまま行われたとしても、食事は人間により提供されたたはずで、当然父親や母親など世話をする人間との限られたコンタクトはあったはずだ。もし動物に育てられた野生児の言語が回復せず、言語を通したコミュニケーションから隔離されたネグレクトの子供の言語は回復するとすれば、希薄であってもこのような人間社会とのコンタクトが特定の時期にあったかどうかが言語の発達可能性を決めている可能性もある。
ろうあ者とSign Language(手話)
既存の言語を介するコミュニケーションは全くなくても、社会の中で他人と関わることで言語が自然に話せるようになることを示す例が知られている。ろうあ者と健常人、あるいは、ろうあ者の間に自然発生する手話だ。現在日本で多くのろうあ者に使われている日本手話(日本語対応手話とは異なる)も、日本語とは無関係にろうあ者のコミュニケーション手段として自然発生してきたものだ。ただ、最初から成り立ちを完全に追いかけることができる手話は多くない。
成り立ちが追跡できる手話は、Village sign language(村落手話:家庭内のHome signも含む)とdeaf community sign language(ろうあ社会手話)に大きく分けることができる。
Village sign language(VSL)は、家族や集落の中で(遺伝的に聴力障害が多い家族や集落である場合が多い)、ろうあ者とコミュニケーションを図るために自然発生してきた手話で、その誕生には必ず健常者が関わっている。中でもよく研究されているのが、4人のろうあの子供を持ったベドウィンの家族内に1930年ごろ誕生し、その後世代を重ねて、4000人の村落のすべてのろうあ者(130人)が共有するようになった手話、Al-Sayyid bedouin sign languagee(ASBL)だ。
ASBLだけでなく、他の自然発生手話からわかることは、発生した手話の文法構造は、家族内の健常人が話している言語の構文構造と必ずしも一致しない点だ。また、VSBLをはじめ様々な自然発生手話同士構文を比べても、特に手話に共通にみられる構文構造は見られない。それぞれのVSLは、それぞれ独自の構造を持っている。このことから、VSLが小さな集団から独自に発生し、発達してきた言語であることがわかる。
現在ABSLを利用している異なる世代のろうあ者が使っている単語や構文ルールが比べられ、最初ほとんど1−2語の構文というより発語から始まったABSLが、世代を重ねるとともに、様々なインプットを統合することで複雑化したことが明らかになっている。最初家族内で使い始めたシンボル化された単語が何で、構文がどんなものだったかについては、調べて見たが残念ながら記載はない。しかし現在では言語と呼ぶにふさわしい十分なボキャブラリーと文法構造を持ったABSLへと発展しているのを見ると、偶然に始まったにしても「シンボル化された単語を他の個体と共有する」スタートが切れれば、参加者が増えるに従って言語が自然に高い機能を持った複雑な言語へと発達することがよくわかる。
しかしABSLが既存の言語の影響を受けずに自然発生したと結論するのはまだ早い。言語を話す健常人は、一つのセンテンスが、各単語を決まった順番に並べること(構文)でできていることを知っており、構文ルールがコミュニケーションに必須であることはよくわかっている。さらにシンボル化された様々な単語を創作する段階でも、健常人の持つ単語の成り立ちや性質についての理解が、手話の成立過程でインプットされた可能性は否定できない。また、健常者も話しながら自然に身振りを使うのが普通で、これが手話発生に影響しないとは言えない。このように、健常者が母国語として使っている言語の構文と異なるというだけでは、VSLを既存の言語から独立した、独自に自然発生した言語であるということは難しい。
ニカラグアの手話
この懸念を完全に払拭し、一定の数のメンバーからなる集団内でコミュニケーションが必要になると、健常者の参加無しに、ろうあ者のみの力で言語を誕生させられることを示したのが、ニカラグア手話だ。
1977年聾唖学校が設立されるまで、ニカラグアのろうあ者の養育は各家庭に任されており、全く手話は存在しなかった。実際、こうして成長した現在65歳以上のニカラグアのろうあ者は全く手話を使うことができない。その後1977年になって、30人規模の聾唖学校が設立され、1979年サンディニスタ政権誕生後は各地からろうあの子供が集められ生徒数が急速に増加、1983年には400人が特別学校で教育を受けるようになった。
この最初に集められた30人の集団から、最初のニカラグア手話が誕生し、これを基礎にして、学校に集まった生徒に受け継がれ、また多くの参加者からの新しいアイデアを取り込んで発展し、現在では子供から大人までニカラグアの多くのろうあ者に使用される言語に発展した。重要なことは、この言語発生過程のほとんどが追跡できることで、最初の発生に関わった第一世代および、その後のボキャブラリーや構文の発展に寄与した後の世代のほとんどが生きており、その時の様子を検証できる点だ。
詳細は省くが、以上の経過から、
図4: 見ると払うの文法的変化を体の向きを変えることで加えている例。このような新しいルールは一旦発生すると、瞬く間に全体で共有されるようになる。(Senghas and Coppola, Children creating language, Psychological Science, 12:323, 2001に掲載されている図。出典:Wikipedia)
例えば図4は Ann Senghasの論文から転載した写真だが、手のサインは「見る」と「払う」を意味している。最初体の中央でだけで提示されていたこのサインは、第二世代になると体の中心だけでなく、左右に振ることで、様々な文法的変化を表すようになっている。しかも、一旦発生して便利だとわかると、瞬く間に同じ言語を使う集団全体に広がることも確認されている。一旦生まれると、言語は急速に進化する!!!
もちろん完全に聴覚を失ったろうあ者の中で自然発生したと言っても、発生に関わった子供達は当然文明の洗礼をうけており、人類最初の言語発生を反映していると結論するのは早いだろう。最初の言語が発生する時にはおそらく、集団で共有できるシンボルを生み出すのには長い時間がかかったかもしれない。しかし、人間は集まると、相手とのコミュニケーションを模索する中で、他の個体と共有できる単語と構文を発生させること、そしてどんなに小さなスタートでも、一旦社会で共有できる言語が生まれると、それ自身が異なる個体からのアイデアを統合して体系的言語へと進化できることを、ニカラグア手話ははっきりと示しているのではないだろうか。
次回は、これまでの議論をもう一度整理し直したいと考えている。
[ 西川 伸一 ]
これまで、言語誕生過程を構想するために7回にわたって言語の重要な特徴について見てきたが、先に進む前にここで少しこれまでのまとめの意味で、情報媒体としての言語の性質を同じ情報媒体のDNAと比較しながら見ることにする。
1、情報は受け手で決まる
現代は情報の時代で、私たちはどこにいっても情報と情報機器に囲まれ、情報なしに生きることはできないと感じている。職場への行き帰りを考えてみよう。時計が指す時刻に合わせて家を出る。駅では次の列車の時間や行き先をみて、遅れや変更がないかを知り、正しい列車に乗る。列車の中を見渡すと、新聞を読んだり、本を読んだり、あるいはタブレットでビデオを見たりしている人もいる。私はといえば、窓の外の景色、時間、車内のアナウンスなどを情報に、目的の駅に降り立つ。駅から職場までの道では、信号の色の指示に従って交差点を渡る。もちろん、信号無視の車が突っ走ってこないかも気をつける。このように私たちの行動のほとんどは、街にあふれる情報に依存して判断していることは間違いない。
半日の行動を思い返して考えてみて、私たちは何が情報で何が情報でないか判断に困ることはない。ところが、私が情報として利用しているものを、抽象的に定義しようとすると簡単ではない。というのも、掲示板の文字も、信号の色も、さらには窓の景色も、すべて職場に通う私には情報になる。一方、視覚障害の方にとっては、窓の景色も、信号の色も、掲示板の文字ですら情報としての価値を持たない。逆に、歩道の凹凸は視覚障害者には重要な情報だが、障害者でないと気にもならない。このように、何が情報かどうかは、それを必要としている受け手に依存している。そして何よりも、世の中のあらゆるものは、受け手によっては情報になりうる。
もちろん受け手だけが情報を決めるのではない。一般的に私たちが情報と認めるものの中には、出し手が情報としての性質を決めているものも多い。例えば、信号や掲示板の情報は、私が利用しなくとも、誰かの情報として提供したいという出し手の意図がある。もちろん、この原稿と同じで、出し手の情報としての意図が実現するかどうかは、受け手にかかっている。いずれにせよ、出し手が決める情報も、受け手が必ず想定されており、情報は受け手で決まる。
これまでの議論をまとめると(人間が受け手の情報に限ってではあるが)、情報は主に「受け手が行動する際に参考にする知識」であり、この中には「受け手の行動を変化させようと出し手が提供する知識」が含まれる。ただ、出し手の存在は必須ではない。突き詰めると、情報とは受け手の行動のための判断に使われる知識と定義していいだろう。
2、情報は物質ではないが、働くために情報媒体(物質)が必要

図1 クロードシャノンと、彼の課題をまとめた図
シャノンは、情報を物質媒体として正確に伝達するために必要な方法(電線でメッセージを伝える電話)を情報理論として体系化した。(写真と図はWiki Commonsより)
次に重要な情報の特徴は、「それ自体は物理的な量ではないが、それが働くためには必ず物理的媒体を必要とすること」をあげることができる。このことを科学的に体系化したのが米国のクロード・シャノンだが、情報媒体が情報と一体化しており、物質からできているので、情報にも物質性があると混同する人が多い。例えば「日本人のDNA」といった使い方がその例だろう。DNAは物質で、情報ではない。「日本人のDNA」の正しい表現は、「日本人のゲノム」だとおもう。
いずれにせよ、情報が物質でないことは少し考えればわかることだ。例えばこの文章はウェッブを通して約4千人の人に読んでもらっているが、読者に内容を紙に印刷して届けようとすると、コストも人手もかかる。これは、情報を媒体に移した途端物理量になり、その結果情報を移動させるのにも大きな物理的力が必要になる。しかしいつものようにウェッブにアップロードすると、一瞬で何千人、何万人、おそらくその気になれば億人単位の人にも同じ情報を送ることができる。これは情報がより分配しやすい媒体(デジタルパケット)に移されたからだが、情報の内容は同じだ。この媒体を選ばない特徴は情報に物質性がないからこそできる芸当だ。
この例では、私が伝えたかった内容(これも私の頭の中の神経ネットワークのパターンとして媒体化されているが)が情報で、PCに私が打ち込んだ時点で情報は電子媒体に移され、液晶画面上の各ピクセルの色の違いのパターンとして表現できるようになっている。このパターンは、そのままウェッブを通して何千、何万のPCに瞬時に伝達できるし、USBでもハードディスクにでも残しておける。
これまで議論してきたのは人間が出し手の(例では私が出し手)情報だが、人間が出し手でない生物が持つ情報、例えば遺伝情報でも同じで、情報自体に物質性はない。またどんなに大きなゲノムでも、DNAやRNAは当然のこと、紙の上のATCGの4文字でも、また電子媒体でも、異なる媒体を使って同じ内容を表現することができる。
もちろん生物の情報には一見物質ではないかと思えるようなものもある。例えばフェロモンにより接合が始まる例では、フェロモンという物質が情報のように見える。しかし、実際にフェロモンが必要とされる状況を考えると、例えば餌が欠乏したという情報をフェロモンが伝えていると考えられることから、フェロモン分子は情報媒体として働いていることになる。
このように、頭の中で情報と情報媒体は混乱し、どうしても媒体を情報そのものと勘違いする。しかし情報は物質でない点を押さえておけば、何が情報で何が情報でないか間違うことはない。人間が出し手の情報は、数学的に記述できても物質ではない。しかし重要なのは、媒体を介して物質世界で働き、物質と相互作用できることで、これこそがシャノンやチューリングにより始まった20世紀の新しい科学革命、情報科学革命だ。
私が頭の中に浮かんだ文章を原稿としてPC上にインプットし、またメモリーにストーレージできるのも、すべてこの情報理論のおかげだ。しかし、人間が出し手でない情報、例えば遺伝情報がどう生物に利用され、脳への情報インプットがどのように記憶として残るのかオペレーション原理についてはわかっていないことも多い。すなわち、情報と媒体との関係は、生物学最大の問題として現在も残っている。
3、情報は生物とともに地球上に生まれた
異論のある人も多いと思うが、私は生命が誕生する以前の地球上に情報は存在しなかったと思っている。しかし待てよ、物理現象を記述するためには、対象となる物質の動きや質量の情報が必須であり、この意味であらゆる分子とその動きには情報が内在しているはずだ。生物誕生以前にも物理現象は地球で続いており、あらゆる物理現象に関して情報も存在していたはずだ。だとすると、情報は生命誕生以前から存在したと言えないだろうか?
答えはNoだ。情報は受け手が決める。物理現象に情報が存在するのは、人間がそれを解釈して利用するときだけで、物理現象が情報を持つかどうかは、それを情報として利用できる受け手の能力にかかっている。人間誕生前にも天体の動きは情報として利用されていたが、それは生命誕生後のことだ。一方、生命自体には誕生時から情報とその媒体が内在していた。この生物だけに情報とその媒体が内在し、また情報の受け手となれるという特徴により、物理世界には存在しなかった全く新しい原理が生まれたことをダーウィンは「種の起源」の最後のセンテンスで美しく表現している。
「もともと生命は、様々な力が、一握りの、あるいはひょっとするとたった一つの原型に吹き込まれて始まり、この惑星が重力法則による永遠に変わることのない回転を繰り返している間に、これほど単純な始まりから、最も美しく素晴らしい果てしない形態が進化し、また進化し続けている。このことを考えると、進化の壮大さに心が打たれる」(私の意訳)
この物理世界にはなかった進化する力を持った生命に最初吹き込まれた様々な力のひとつが、情報とその媒体であることは間違いない。
生命誕生を情報の誕生として認めると、地球最初の安定した情報媒体は核酸ということになる。では、この核酸が媒介している情報とは何か?答えは、ATCG4塩基の配列(コード)になるが、実際に核酸が媒介している情報はこれに止まらない。その先には、塩基の配列によって決まるRNAやDNAの構造や機能、アミノ酸に翻訳される場合はアミノ酸の配列、アミノ酸が形成する分子構造とその機能、さらには、その分子と結合する様々な分子、などなど情報のコンテンツは拡大する。わかりやすい例で言えば、「空」という単語の中に、太陽、月、星、そして無限の宇宙は言うに及ばず、それと反対の概念である陸までもが収束しているようなものだ。
このような媒体に無限の情報が表現されているという特徴は、生命の情報媒体全てに言えることではない。例えばフェロモンが担える情報には限界がある。しかし、核酸、脳回路、そして言語の3媒体には、このような限界は全くない。これまで見てきたように、3つの媒体とも質・量ともに無限に拡大できる。また、媒体をコピーすることができる(正確である必要はない:文字が生まれるまで言語も決して正確にはコピーできない)。さらにその媒体が表現している情報と外界との相互作用を様々なレベルで記録することができる。この特徴のおかげで、生命や人間の進化は進んできた。
4、DNAと言語の比較
DNAは情報媒体として働くために、4塩基配列をコードとして使っている。このコード自体は物質ではなく情報だが、媒体と一体化しているので、情報と実際の世界をつなぐ接点として考えればいいだろう。では、言語で塩基配列に対応するコードはなんだろう?これは人間が発声できる音の配列と言える。
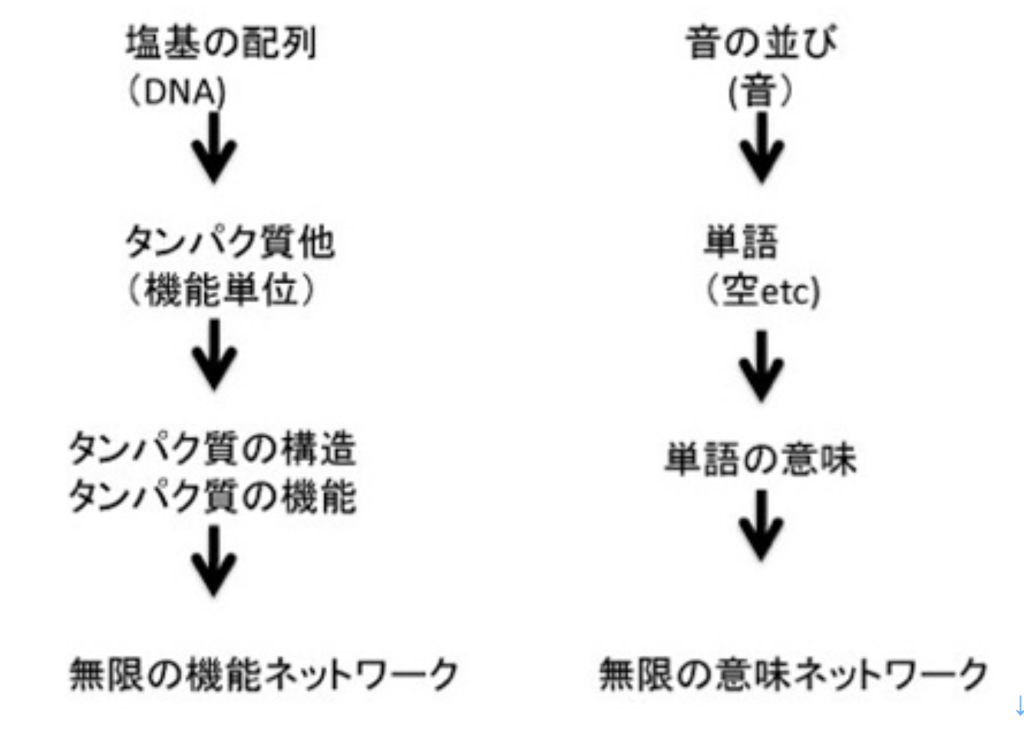
図2 言語とDNAを媒体とした情報の比較
次にコードにより表現された単位が来る。言語の場合は単語だが、DNAの場合はアミノ酸からなるタンパク質だけではない。そこでとりあえず機能単位としておく。ただ、話を複雑にしないため、これ以降はアミノ酸に翻訳されてできるタンパク質が、言語での単語に相当するとして話を進める。
ここまで、なんとなく言語とDNAは似ているように述べてきたが、一つ大きな違いがある。すなわち情報の媒体の物質性だ。DNAはタンパク質に対応する情報を表現する媒体だが、この延長で言えば言語を媒介する音は情報の媒体であるのは間違いないが、物質性が希薄で、かなり特殊な媒体であるといえる。
例えば、文字が誕生するまで、言語の記録は私たちの脳内神経ネットワーク以外に記録することは難しかった。要するに、言語の体系は覚える以外、維持することは不可能だった。当然脳の記憶の仕組みから考えると、記録という面ではなはだ心もとない。しかし、以前議論したように言語の場合、個人の脳内に維持されている表象とともに、集団で共有される部分を持つ2重構造になっている。
言語の集団共通部分ではさらに物質性が希薄になる。しかし、集団で共有しているという性格のおかげで、各個人の脳内神経ネットワークの記憶の限界を補うことが可能になっている(次回もう一度議論する)。このように、文字が生まれる前の言語は、音という物質性の希薄な媒体(ほとんど持続できない)に依存しているため、この物質性のなさが、DNAとは大きく異なっていると言える。ただ、この問題は言語の2重構造を実現することで、ある程度解決している。幸い、言語媒体の抱える物質性の希薄性の問題は文字の誕生で解消する。
5、DNAも言語も、媒介する情報は部分と全体が常に一体化している。

図3 メタボリックマップと、分子ネットワーク
もともと生物学では、一つの遺伝子にコードされている分子も、大きなネットワークの中で様々な分子と関係を持って存在するとして研究が行われてきた。例えば、ある分子が加わる代謝マップに存在しないということすら情報になる。(Wiki Commonsより)
図3に代謝マップと、分子ネットワークの例を示したが、情報媒体としてのDNAから学ぶことができるもう一つの重要な点は、一つの情報は決してそれ自身で存在していないことだ。これまで、生命の情報を扱ってきた分子生物学は、どうしても個々の遺伝子に焦点を絞って研究してきた。このため、ともするとDNA媒体により表現されている情報が、特定のタンパク質についての情報であると思ってしまう。しかし、研究対象に選んだ遺伝子が一種類のタンパク質だけをコードしているとしても、その分子は生物の中で他の分子と直接結合するだけでなく、多くの分子と直接間接に相互作用を行っている。すなわち、一つの分子の情報の背景にはその分子だけでなく、多くの過程が背景として存在している。更にある細胞にこの分子が発現しているということだけではなく、例えばアルブミン分子をコードする情報には、赤血球や多くの細胞にはアルブミンが存在しないということも含まれている。結局個々の情報も、全てのゲノム情報と関連している。このことは生物学者なら肝に銘じていることで、生物のもつ部分と全体の特別な関係として研究者の頭を悩ませてきた。
しかし、言語になると、多くの人が言語体系は単語という部分が単純に集まったものとして考えてしまう。これは、多くの人が、言語とは意味しているもの(シニファン:記号)と意味されるもの(シニフェ)が一体化した記号が集まったものだと述べたソシュールの考えを鵜呑みにしているからかもしれない。しかし実際に私たちが「空」という言葉を頭に浮かべる時、決して大きな広い空だけではなく、太陽、月、星、海、陸地など同時に多くの単語を思い浮かべるのが普通だ。実際には空と聞いて、星を思い浮かべていることもあるだろうし、水平線を思い浮かべていることことすらあるだろう。このように、私たちの厖大な経験の一部を単語として記号化したのが言語で、決して記号を集めて経験を作り直しているわけではない。すなわち言語はシニフェとシニファンが一体化した記号を集めて形成されたものではない。
この点を理解するには、言語ともう一つの情報媒体、脳の神経ネットワークとの関係を見る必要がある。次回はこの問題を「言語と脳ネットワークを媒体とする情報の比較」というタイトルで考える。
[ 西川 伸一 ]
前回DNAなど核酸と言語について、情報媒体としての観点から比較して、それぞれはあらゆる情報を表象することができるものの、言語を媒体として使う時に必要な「音」がDNAと比べた時、物理性に乏しいことを強調した。もちろん音も物理現象で、物理現象だからこそ情報を媒介できる。誤解を招かないためには、本当は音が物理性に乏しいという代わりに、音が持続性に乏しいといったほうがよかった。しかし、DNAが生物の情報媒体として誕生して以来、新しく進化してきた情報媒体はほとんど持続性に欠ける媒体で、決して世代を超えて情報を伝えることはできなかった。その最たるものが、情報の媒体としての脳の神経回路だ。事実、ある時自分が見たり考えたりしたことを覚えておくのは難しいし、ましてや他人や子孫に伝えることはもっと難しい。脳の神経回路を媒体とする情報の延長上に発展してきた言語が持続性に欠けるのは当然と言っていい。そこで今回から2回に分けて、情報媒体としての神経回路を言語を念頭に整理し直して、言語が神経回路を媒体にした情報処理システムに何を新たにもたらしたのか考えていきたい。
本題に入る前に、情報と情報媒体にまつわる混乱を整理する意味で、「言語は媒体か?情報か?」について少しだけ考えてみよう。言語は私たちが感じたり、考えたりした内容(=情報)を表現するための媒体として利用されている。ただ、情報を表現するためにはどうしても、単語やさらに小さな単位の音節を一定の法則(文法)に従って並べる必要がある。この音節(単語)の並びに情報性が生まれるのは当然で、並びを正確に伝達する方法の開発が、シャノンの情報科学が生まれる発端となった。同じことは、DNAにも言える。DNA自体は情報媒体だが、一本のDNA鎖は異なる領域に分節され、それぞれの分節は4塩基の配列として表現されている。従って当然配列自体が情報性を持つ。このように情報媒体は情報を背負った途端にそれ自身が情報になる。これが情報と、情報媒体の区別についての混乱の原因になるので注意が必要だ。ただ、音の並びや、DNAの並びを情報として捉えて混乱させているのは、わたしたち人間自身で、実際には言語も、DNAも情報媒体以外の何物でもない。
本題に移ろう。脳については、神経細胞の進化過程から人間に特有な高次機能まで、何回にもわたってその情報処理の仕組みについて説明してきた。これまで議論した詳細については理解していただいていると思うが、おさらいをかねて、まず私たち人間の脳について、情報媒体という観点から整理してみよう。
さて今年のノーベル賞は概日周期のメカニズムを発見した3研究者に与えられたが、情報とは何かを考える良い材料になるので、概日リズムの話から始めよう。
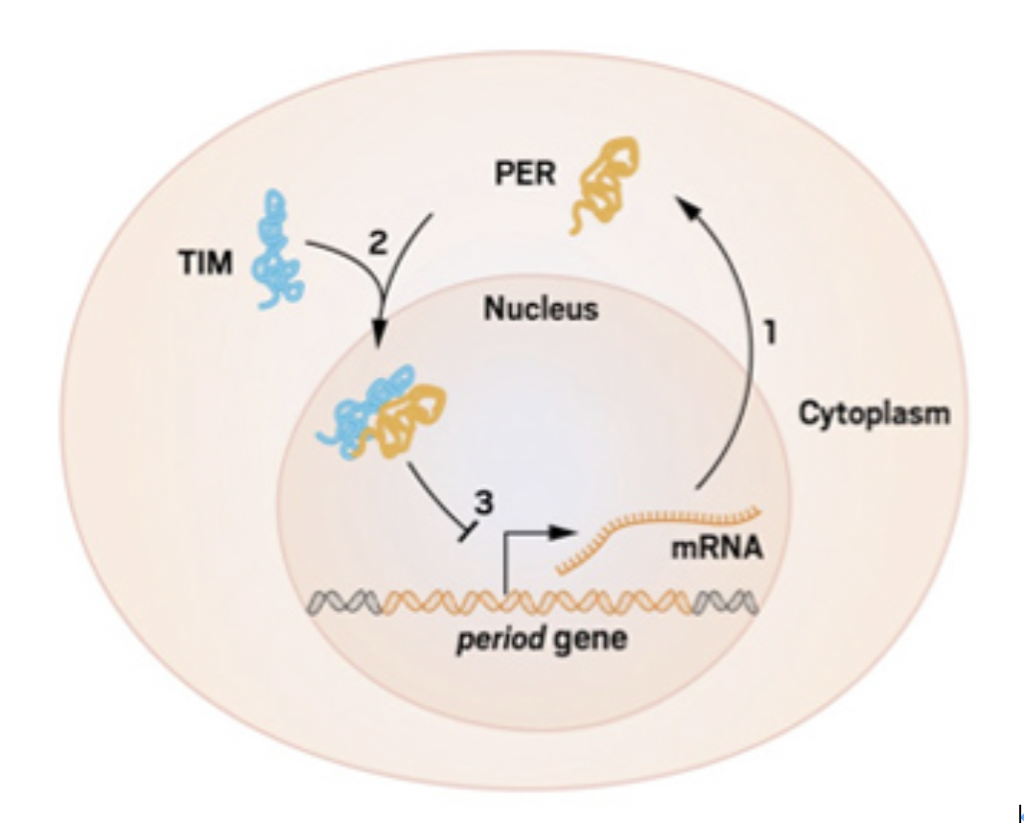
図1:ノーベル財団の今年の医学生理学賞の受賞理由に掲載されている図。
(https://www.nobelprize.org/nobel_prizes/medicine/laureates/2017/press.html) (C) The Nobel Assembly at Karolinska Institute
「DNAは情報媒体として、概日リズム、すなわち地球の自転情報をコードしている」というと、驚かれるかもしれない。図1はノーベル財団から発表された受賞理由に掲載されていた図だが、概日リズムのメカニズムを説明している。しかし目を凝らしても、この図には地球の自転を感知する仕組みは一切見当たらない。それもそのはず、私たちの体にある一つ一つの細胞が概日周期を持っており、これが全てゲノムにコードされていることを明らかにしたことが今回のノーベル賞の受賞理由だ。この細胞レベルの概日周期も、視覚を通して感じた地球の自転情報で調整し直すことが可能だが、細胞の概日リズムの維持に光を感じることは必須ではない。これは、体から分離した培養細胞を真っ暗な部屋で培養しても、リズムは維持されることからわかる。このリズムは、生物進化の過程で、地球の自転という情報がDNAを媒体とした情報としてゲノム上に書き込まれた結果だ。
このことから、地球の自転のような宇宙レベルの情報ですらDNAを媒体とした情報へと書き換えられること、すなわちDNAがほぼ無限の情報を媒介できることがわかる。ただ、DNAに地球の自転情報を書き込むためには、偶然によるDNAにコードされた情報の変化と、変異した情報から生まれる形質の変化を選択し、最も外界にフィットした情報を固定化する途方もない進化の時間が必要だった。このことからDNAを媒体とする情報は、外界を記憶するというより、外界の情報(この場合地球の自転のサイクル)を自己に同化していると考えるほうがいい。しかし組み込む過程は不自由で機動性に欠けていても、DNAを情報媒体としてこれまで生物ゲノムに同化された外界の情報はほぼ無限と言ってもいい。
繰り返すが、この同化過程は、情報媒体に起こる偶然の変化に依存しており、外界の変化に素早く反応することは全くできない。例えば、個体の一生と言う時間スケールで情報に合わせて変化したり、あるいは情報を他の個体に伝達することは不可能だ。このことは、大腸菌を特定の栄養成分が欠けた培地に移して変化に適応させる時、ほとんどの個体が死滅する中で、何百万分の1の確率で生き残った個体だけが、次の世代を作るのを見ればよく理解できる。しかし、個体が外界の変化に適応して生きるためには、素早く外界の変化に適応できる能力の開発が必要なのは明白だ。この問題の解決として、様々なシグナルを使って外界をモニターする方法(例えばクオラムセンシングなど:2016年8月15日 進化研究を覗く)が進化した。ただ、情報の解釈、及び記録の両方を実現できる情報媒体として進化したのは、ヒストンやDNAの修飾により遺伝子の利用を決めるエピジェネティック機構だ。この機構の進化により、外界の変化を受けて、細胞を安定的に外界に適応した状態へとシフトさせることが可能になった。この結果、細胞同士がコミュニケーションすることでそれぞれの分化状態を決めることが要求される多細胞体制も可能になった。
エピジェネティック機構はDNAの書き換えを必要としない。その代わりに、細胞内外の情報に従ってDNA上のヒストンやDNA自身を可逆的に修飾し、遺伝子発現を変化させる。すなわち、ゲノム情報の中の特定の組み合わせだけを機動的にON/OFFできるようにすることで、一種の記憶を可能にしたと言える。実際、原則としてエピジェネティックな情報は次世代に伝達されない。しかし、同じゲノムを共有する個体であれば、同じ外界からの刺激をエピジェネティックな記憶として多くの個体で共有することができる。言い換えると、生殖を通してしか伝達できないゲノム情報を、何通りにも違った使い方をできるようにしたのがエピジェネティックな機構と言える。
ただ、エピジェネティック機構を用いた媒体での情報処理には時間がかかり、迅速性にかける。一方、生物はさらに短い時間スケールの変化に囲まれている。特に、光、音、温度のような物理学的変化は短い時間スケールで変化することが普通だ。そして、これらの変化は生命にかかわることもある。しかし、神経系が発達するまでこのような早い変化に生物はついていくことができなかった。言い換えると、これらの問題の解決として進化したのが神経系と言える。
イオンチャンネルを通るイオンの流れをシグナルとして使うことは神経細胞以前から行われている。分かりやすいのが、ミトコンドリアのATP合成に使われるチャンネルだろう。しかし、電位差によって開閉するチャンネルを使って、細胞の端から端まで順番に活動を伝播させるシステムは神経系が最初だ。この膜電位の脱分極による興奮性は、急に開いたチャンネルを通るイオンの流れなので、刺激に対する秒単位の反応が可能になった。
図2:神経系はすべてのシグナルを、膜の興奮に収束させている
これに加えて、刺激の性質の異なる外界の情報を、細胞膜の興奮という統一した仕組みにいったん収束させることが可能になったことが(図2)、情報媒体としての神経系の特徴だ。この結果、神経系では、物理刺激も、化学刺激も全て同じ興奮原理を持った神経回路に統合して統一された情報として扱うことが可能になった。これにより、あらゆる種類の外界の変化を、迅速に捉え、一つの記憶として維持することができる、まったく新しい情報媒体が出来上がった。
ただ、膜の興奮そのものは、記憶というより、反射に近い。神経細胞内では、この迅速な反応は、短期、長期記憶と呼ばれる二つのメカニズムで持続される(図3)。
図3:短期記憶と長期記憶のメカニズム(ノーベル財団2000年ノーベル医学生理学受賞理由:https://www.nobelprize.org/nobel_prizes/medicine/laureates/2000/press.html) (C) The Nobel Assembly at Karolinska Institute
図3は、エリック・カンデルの研究を紹介したノーベル委員会の受賞理由に掲載されたいた図を拝借したものだ。この図には、神経細胞内でのシグナル伝達系(PKA, cAMPなど)の活性による短期記憶と、エピジェネティックな変化によって誘導される細胞の分化を用いた長期記憶のメカニズムが書かれている。すなわち、神経系の誕生で、これまでシグナル伝達や、細胞分化に関わってきた多くのメカニズムが、新しく誕生した興奮膜を中心に再編成されているのがわかる。
図4:情報媒体の進化の階層性
このように、DNA、エピジェネティック機構、神経と情報媒体の進化を振り返ってみると、新しい媒体が、古い媒体では困難だった様々な課題を解決するとともに、それ以前の情報媒体を、新しい媒体を核に再編成し直して利用していることがよくわかる(図4)。この結果、神経系では、興奮膜の反応時間から、エピジェネティック機構の時間、さらにはゲノムの時間まで統合された独特の時間を形成するのに成功している。
これらの時間は、一個の細胞内の時間で、例えば以前紹介したゴカイ幼生の神経が一本しかない光感受システムにも当てはまることだが、記憶・伝達という観点から見た時、神経系は別の方法でもうひとつの時間過程を形成するのに成功している。すなわち、シナプス形成による神経細胞同士が興奮を伝達できる回路の形成だ。例えばアメフラシの水管反射回路(図5)を思い出してもらいたい。この回路で神経細胞はシナプスで結合している。個々の細胞は刺激に応じて長短合わせた変化を遂げシナプスの興奮性も変化するが、これとは別に回路内では一つの細胞から細胞へと刺激のリレーが行われ、この興奮のリレー自体も回路特有の活動時間を形成している。すなわち、外界からの刺激に対して、細胞やシナプスといった個々のレベルだけでなく、回路全体として反応が起こり、記憶が形成される。
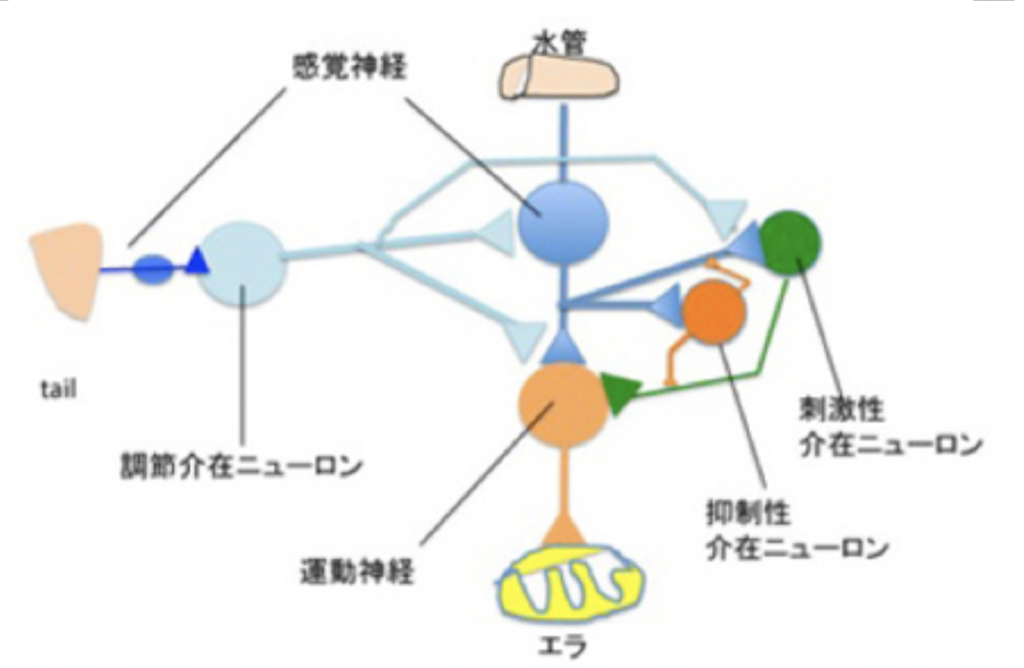
図5 単純なアメフラシの水管反射に関わる回路。(BRHホームページ参照:http://www.brh.co.jp/communication/shinka/2016/post_000024.html)
そして、この細胞間のつながりは、ほぼ無限に拡大できることから、私たちが例えばイメージを見たとき、脳の中では恐ろしく複雑な過程が進行し場合により記憶として回路レベル、細胞レベルに分散して残ることが推察できる。
この外界の刺激を認識、記憶する過程の延長に、言語が生まれるのだが、例えば、鳥を見たとき脳で起こっている過程の複雑さを考えると、「トリ」という言葉が刺激として持つ性質は、あまりにも単純に思える。私自身は、このギャップこそが、言語の重要な役割だと思っているので、次回は、さらに高等動物の記憶について整理しながら、言語の役割について考えてみたい。
[ 西川 伸一 ]
DNAから神経系まで、生物の持つ情報媒体をざっとおさらいしてきたが、今回は高次脳機能としての記憶について焦点を当て、言語との関係で考えてみたい。前回見たように、新しい情報媒体は、それまでの媒体ではできなかったことを実現してきた。とすると、言語は高次の脳機能ではできないことを可能にしたことになる。ただ、言語によって何が可能になったのかについて理解するためには、言語発生直前の脳を知る必要があるが、簡単ではない。結局現在の私たちの基本的な脳機能を理解した上で、何が新たに可能になったのかについて想像をめぐらすしかない。いずれにせよ、エピゲノムがゲノムの延長にあるように、言語は脳の高次機能の延長上に生まれた。個人的には、言語は動物の脳の持つ記憶とコミュニケーション機能の新しい展開として誕生したと思っているので、言語を念頭に置きながら、高次の記憶やコミュニケーション能力について考えてみよう。
記憶については既に何回か折に触れて説明してきたが、私たちが記憶として通常思い浮かべるような複雑な記憶について説明はしてこなかった。この問題が現在も研究途上で説明が難しいためだが、今回はあまり詳細にこだわらず、独断で記憶を論じてみたいと思う。
図1 細胞の短期変化から細胞の安定な分化が細胞レベルで記憶を支える。前回の図3も参照
まず記憶の細胞学的メカニズムを復習しよう。記憶は、神経細胞自身の興奮伝達特性の変化と、その結果起こる神経間の結合性の変化、そしてそれに続く神経ネットワークの変化により支えられている。前回再掲したエリック・カンデルのノーベル賞受賞業績紹介に掲載された図をもとに、記憶の持続時間という点から書き直してみると図1のようになるだろう。
まず神経刺激により誘導される生化学変化により短期間神経細胞やシナプスの刺激反応性が変化する。この刺激が一定期間続くと、次に新しい遺伝子発現が誘導され、シナプスの興奮伝達に関わる分子がさらに長期的に変化し、ポジティブ、ネガティブに伝達性が変わる。さらにこの過程が最終的にエピジェネティックな変化を起こすと、新しい神経間の伝達性の変化を安定的に維持することができる。この中には、シナプスの消失や、形態変化(例えばスパインの変化:http://www.brh.co.jp/communication/shinka/2017/post_000005.html)、さらには細胞の増殖による新しいネットワークの組み替えも含まれている。神経細胞は多様化しており、シナプス形成様式も多様で、個々の神経レベルではこの過程に様々な分子が関わるが、基本的なメカニズムは同じだ。
ただ細胞レベルのメカニズムからだけでは私たちの記憶システムを説明できない。これは、複雑な記憶には脳神経回路によるプロセッシングが必要で細胞レベルのメカニズムが共通だとしても、多数の細胞が参加する回路ができることを抜きにして、高次な記憶は形成できない。この時、外界のイメージから感覚器を介して入ってくる脳へのインプットを、神経回路活動の様々なパターンへと分解し、この中から必要な情報を集めて、知覚したイメージに対応する内部イメージが神経回路内に作り直される(=すなわち外部のイメージが脳内に表象される)。この表象過程では、表象の再構成に必要な断片化された情報を集め直す必要があるが、そのためにそれぞれの情報を神経ネットワークに一時的に維持しておかないと、統合することは難しい。これらを短期の作業記憶と呼ぶが、新しい表象は短期に記憶された情報断片の統合として現れる。こうして生まれた新しい表象の中から、さらに長期に記憶する表象が選ばれ、私たちの記憶が誕生する。
この過程をもう少しわかりやすく説明するため、多くの絵画が展示されている、美術館の常設展示場を訪れたという状況を考えてみよう。私の場合、一枚ずつ足を止めて絵を見、またラベルから絵のタイトルや画家の名前を丹念に拾っているのだが、一度見ただけでは絵の詳細について覚えられない。したがって、絵を見て脳内に形成されたほとんどの表象は時間とともに消えてしまう。しかし、5年後にもう一度同じ展示場を訪れた時、絵が以前と変わりなく展示されていたら、確かに前に見たと思い出す絵は多い。これは、絵の表象を覚えていたというより、部屋の雰囲気などを覚えており、もう一度絵を前にして、初めて見た時の感覚が蘇ってくるのだろう。さらに、前もって美術館の予習をしておけば、絵を覚えられる可能性は高いし、画家について知識があるさらに覚えやすい。もしその画家が自分の好きな画家なら、もう忘れない。最近の研究では、スマフォで写真を撮るだけで、後で見直さなくても絵を覚える確率は高いようだ。要するに、絵を見るだけでなく、様々な情報が合わさると記憶に残るようだ。
しかし、知っている画家の絵だけが記憶に残るわけではない。それまで聞いたこともない画家の絵に強い印象を受けて忘れられなくなる絵も多い。個人的経験だが、アントワープ王立美術館で見たフーケの聖母子像がそうだった。それまでフーケの名前を全く知らなかったが、その時以来40年経っても忘れることはない。(余談だが、他にもJan Gossaertの艶かしい絵をはじめとして、個人的にはフランドルの画家の絵は一度見ただけで忘れられない絵が多かった。)

図2 アントワープ王立美術館所蔵のジャン・フーケの「ムランの聖母子像」、1981年に一度訪れた後は訪れる機会がないが、イメージは鮮明に残っている。
(出典:Wikipedia)
このような美術館での体験の記憶は短期、長期を問わずエピソード記憶と呼ばれる。
フーケの絵(図2)の前に立ったと想像してみよう。私たちは、決して写真機のように絵全体を認識しているわけではない。特に近くから見るときは、ほとんど無意識に(おそらくこれまで培ってきた習性に従って)、例えば顔や、乳房(私の場合だが)などの部分部分に視線を走らせ、イメージを取り込んでいく。このとき、網膜の特定の場所で補足されたイメージは色と形に別々に処理される。また無意識に視線を動かしていても、常にこの動きは視覚とは別にモニターされ、立体感を得る時の情報として使われる。要するに、一つのイメージはバラバラの要素として脳にインプットされるため、それぞれを一定時間保持しないと、全体を再構成しなおすことはできない。この各要素を短期の記憶として維持し、統合する過程に必要なのが「作業記憶」で、図3に示すように海馬と視覚野に関わる脳の4−5領域がネットワークを形成してこれに当たっている。この中で海馬が最も重要な役割を演じており、海馬が障害されると、新しい記憶を成立させることができなくなる。
図3: 視覚の表象形成と作業記憶。
網膜に結像したイメージは後頭葉の視覚野に投射されるが、同時に動眼筋肉など様々な視覚に関わる情報が脳内の様々な領域で処理される。こうして集まった情報は、海馬を中心に一時的に記憶される(作業記憶)。もし情報が足りない場合、ネットワークからの刺激により、足りない情報が集められることもあり、表象を完成させるためには、海馬を中心として様々な領域が活動する。こうしてできた表象は、さらに多くの脳領域と連合される。
このように視覚から表象が形成される過程に限ってもこれだけ複雑で、美術館ではそれぞれの絵を見ながらこの過程を繰り返すことになる。もちろん、絵を見ている時、聴覚を始め様々な感覚も並行して脳に入ってくる。それ以外にも、絵を見ながら様々なことを考えることもある。この場合、絵の表象を短期的に形成する過程が、他の脳領域の活動の影響を受けることになる。音が気になりすぎたり、絵とは関係ないことを考えたりしてしまうと、絵の表象の形成は抑制され、絵を見たことさえ気がつかないことすらある。しかし、同時に聞こえる音や匂いで、絵の印象が余計に強くなることもある。ふっと浮かんだ考えが、絵のイメージを強くすることもある。
神経興奮の基本メカニズムはあらゆる細胞で共通だ。絵を見ることも、作業記憶も、表象の形成も、そして連合も全て、この共通の神経興奮メカニズムを基盤にしている。そのおかげで、一つの神経活動は、他のあらゆる活動と相互作用することができる。その結果が記憶に影響して何の不思議もない。これが、記憶の連合と呼ばれる現象だ。
知っている画家の絵の方が覚えやすいのは、内部イメージとして存在する画家の知識が、今見ている絵の表象と脳内で連合されるからだ。画家を知っているという知識がどのように脳内で保持されているのか詳細はわからないが、今出来たばかりの絵の表象とは違い、年月を経た安定した長期記憶だ。新しい表象も安定した表象と連合することで、安定な記憶へと変換できる。
面白いのは、例えばスマフォで写真を撮るだけで絵が覚えやすくなるのは、スマフォで写真を撮影したという単純な記憶が、絵を見たという複雑な記憶と連合することで起こると推測される。2度目に展示室を訪れて、絵を思い出すのも、部屋の雰囲気といったより単純な記憶が絵と連合しているからではないだろうか。すなわち、単純な表象と連合させることで、複雑な表象が覚えやすくなる。この記憶の特徴は言語の発生を考える時に鍵になる。
一方、私が初めてフーケの絵を見ただけで記憶できたのは、情動が新しい表象の連合を後押ししてくれたおかげだ。すでに述べたが(http://www.brh.co.jp/communication/shinka/2017/post_000010.html)、情動は私たちの行動のエネルギー(力動)として、脳活動全体を支配する。情動を引き起こすためには、これまでの私の好みなどについての記憶が必要だが、強い情動が生じると、情動自体と、あるいは他の表象との連合の程度が高まり、2度と忘れることのない絵の記憶が私の脳の中に成立する。恐怖体験や大きな喜びの体験はよく覚えているのも同じことだ。
以前、刻々と知覚される膨大な情報が、神経ネットワーク上に形成された自己というフィルターで選択され記憶されることで、新しい自己が脳内にできることについて述べたが(http://www.brh.co.jp/communication/shinka/2017/post_000007.html)、こうして考えると、美術館での体験による表象の形成、連合、そして記憶と続く過程は、この新しい自己を脳神経ネットワークに書き換えることと同じであることがわかる。異論もあると思うが、私は長期記憶とは脳神経ネットワーク上の自己が新しく書き換えられたことだと思っている。
美術館での絵の鑑賞を例に記憶の成立に至る脳過程を見てきたが、短期の作業記憶から表象が形成され、最後に長期記憶として自己の脳神経ネットワークに統合される過程が、鑑賞している一枚一枚の絵で繰り返されるとすると、ほんの一部を除いてほとんどの絵の詳細は美術館を出た時には忘れているのも当然だ。実際、私たちが持つ作業記憶のキャパシティーは少ないと考えられている。アルツハイマー病で最も問題になるのがこのキャパシティーの少ない作業記憶だ。
しかし、この作業記憶が長期記憶に移行するときは、すべての作業記憶が記憶されるのではなく、より単純化された情報量を持つ表象が記憶されるのだと思う。情報量は低くとも、一つの表象が単独で記憶されるのではなく、それまで蓄積した全表象とつながるネットワークの一部として記憶されることで十分記憶として成立する。この一つの表象が自己の脳神経ネットワークに統合され全体と関わることも、言語の発生を考える上で重要なポイントになっている(最後に議論)。
では、40年後の今、私がフーケの絵を思い出す時、脳内で何が起こっているだろう?目を閉じて絵を思い浮かべてもらいながら脳のどこが活動するかを調べると、なんと網膜から直接投射を受けている後頭葉にある一次視覚野が興奮することがわかっている。この結果は、脳内に記憶している内的イメージを思い浮かべる時、私たちは記憶から要素を選び出し、これを一次視覚野で集めなおして、あたかも新しい網膜刺激を受けたように思い出していることを意味する。とは言っても、今見てもらったばかりのフーケの絵を目を閉じて思い出す時、決して写真と同じように思い出していないはずだ。実際、赤い天使の中に浮き上がった真っ白い聖母が、左の乳房をキリストに与えようとしているといった大まかな像が浮かんでくる。しかし、夢を見るように鮮明なイメージがそのまま浮かぶことはないと思う(少なくとも私は)。写真を見直すと、鮮明なイメージを覚えているように思ってしまうが、実際には記憶されている内部イメージは簡略化されている。
記憶の成立から、呼び起こしまで詳細を省いて概略を述べてきた。この過程からわかるように、鮮明な記憶を維持することは膨大な神経活動が関わる大仕事で、毎日の一瞬一瞬を覚えることなど不可能に近い。代わりに、生まれてから神経ネットワーク状に形成してきた自己をフィルターとして、作業記憶から現れる表象の一部を随分簡略化して自己のネットワークに統合することで、長期記憶を成立させている。これが簡略化されていることは、成立した内部イメージを呼び起こそうとするとわかる。脳内からそれぞれの情報が一次感覚野に集められ、感覚として再現される。ただ、視覚のように複雑なイメージの場合、鮮明に思い出すことは難しいことから、決して写真のような鮮明なイメージ全体を記憶しているわけでないことがわかる。
このように記憶を理解すると、記憶にとっての言語の意味、すなわち言語にしかできないことが見えてくるので、最後にそれをまとめておく(検証された概念ではなく、私の個人的な妄想と理解してほしい)
まず、強い感情、絵が展示されている部屋のイメージ、あるいはスマフォで写真を撮ったという単純な行為と連合させるだけで、複雑な絵が覚えやすい例からわかるように、複雑な表象も感情や他の単純な表象と連合することで、覚えやすくなる。とすると、音の短い並びからできた情報量の少ない単語は、連合させることで複雑なイメージの記憶を助ける力がある。確かに、私たちの記憶はかなり言語に助けられている。言語を持たない動物の長期記憶のレパートリーは、私たちよりはるかに感情との連合に頼っているのではないだろうか。
言語(単語)は情報量としては単純なおかげで、記憶するために連合させる相手としては優れているため、最初は各表象が自己の神経回路に統合される助けとして脳内神経回路の一部として取り込まれる。重要なことは、脳内回路に取り込まれるということは、単語が一つのイメージに対応した表象として独立に存在するのではなく、自己の脳神経ネットワークに組み込まれた他の多くの表象と関わって存在していることを意味する。とすると、当然言語は「リンゴ」がリンゴにだけ対応するのではなく、リンゴではないミカンや、ブドウとも関わって、記憶を助ける働きを発揮する。この一つの単語が最初から、多くの他の単語と関わるという構造が、言語の最も重要な特徴になる。
最後に、内部イメージを呼び起こそうとしても、鮮明なイメージを呼び起こすのは難しいことを考えて欲しい。不明瞭なイメージでも、同じイメージに再び出会った時に、「あいつだ」と思い出すためには十分役にたつ。しかし、頭の中でプランを練るといった場合にイメージをこれ以上鮮明にするのは難しい。しかし一度フーケの絵を思い起こす時、「赤い天使」、「白い肌のマリア」、「腕の中のキリスト」、「キリストに飲ませる左の乳房」、と言葉を同時に呼び起こしてみると、イメージは格段と鮮明になる。これが、すなわち記憶の質を大きく高めてくれる。
最後に、今回議論した記憶は、陳述記憶や意味記憶と呼ばれる、脳神経ネットワークの連合を広げる方向での記憶だが、逆に回路を限定することで形成される記憶も存在する。例えば、一度自転車に乗れるようになると、もう乗り方を忘れないような、いわゆる熟練と言われる記憶で、手続き記憶と呼ばれている。個人的考えだが、言語には、この手続き記憶も重要な働きをしていると思っている。すなわち、単語と対応する最も重要な表象を最短で繋いで、意味記憶システムの関与を整理している。記憶という観点から見ると、言語は意味記憶と手続き記憶が統合された脳活動ではないかと思っている。
しかし、これらはすべて記憶に対する言語の意味で、記憶システムが言語のような情報量の少ない連合の相手が必要としたとはいえ、この要求だけから言語が生まれることはないと思う。そこで、次回はコミュニケーションについて考えてみる。
[ 西川 伸一 ]
前回、記憶の機能がいかに言語により高まったかについて私の考えを説明したが、駆け足だったので分かりにくかったかもしれない。そこで、図を使ってもう少し記憶と言語について説明してからコミュニケーションの問題に進むことにする。
私たちの脳への入力情報の7割以上が視覚からの入力であり、前回述べたが視覚表象の形成やその記憶は途方もなく複雑だ。ただ、作業記憶や長期記憶を形成するとき、より単純な情報と連合させることで、記憶を高めることができる。その最たる例が感情との連合で、おそらく言語を持たない動物の記憶の大半は感情と連合しているのではないだろうか(満開の花の思い出も、団子と連合している点で人間も同じだが)。この最も分かりやすい例が、メトロノームの音で、食べ物を連想させるパブロフの反射だ。同じことを、たまたま存在する簡単な表象と連合させることで記憶力を上げることができるが(mnemonicと呼んでいる)、都合の良い単純な連合相手はそうあるわけではない。この連合相手の機能を持つのが言語で、それ自身は情報量の少ない音の並びからなる単語を脳内で表象して連合の相手とすることで、複雑な情報の記憶を高めることができる。
言語を記憶のためのmnemonicな印(一種メモと考えればいい)と考えると、単語がそれに対応する対象の表象とリンクした途端、その表象は他の様々な表象が含まれた脳内ネットワークの中に組み込まれることになる。言語が存在する前から、りんごの表象は、ミカンやブドウの表象ともネットワークを作っており、さらには果物というカテゴリーの表象とも連合している。従って、単語の表象が実物の表象と連合した時から脳内の様々な表象のネットワーク内に言語も体系化されることになる(図1)。
とはいえ、りんごそのものの表象と、「りんご」という単語の表象は、ネットワークの中では、反射的に双方が想起されるほど密接に結びついている。これは音と実物の表象をリンクさせるパブロフの条件反射と同じで、この連合は手続き記憶成立に似た過程で形成されるのだろう。この結果、無意識にりんごという単語からりんごが浮かぶようになる(黄色の矢印)。さらに、果物は例えば食べると甘くて美味しいという感情に裏付けられた行為の記憶によってカテゴリー化されているだろう(赤い矢印)。すなわち、りんごの表象がリンクしている一連のアクションを表象するネットワークに単語もリンクすることになり、このアクションの表象が「私・食べる・りんご」といったプリミティブな文法の元になるように思える。
図1:メモ代わりの標識表象としての言語が脳内の表象のネットワークに組み込まれて、言語が成立する。言語としての統語は、それぞれの表象が属する様々なカテゴリーによって決まる。例えば、食べるという表象が最初から果物には属している。このような言語の構造のおかげで、新しい単語を学ぶとそれは実物についての表象を介して脳全体の表象ネットワークの中に位置付けられる。これが、私たちが急速に単語を覚えることのできる基盤だと私は思っている。
図1を見ながら繰り返すと、(文字のできる前の)言語とは、脳内に形成した重要な表象を覚えておくための単純なmnemonic表象として脳内ネットワークに形成される。脳内の複雑な表象と、単語というそれ自身音の並び以外の意味を持たない単純な表象(シンボル)が脳内で手続き記憶を通して実物の表象と結合した途端、単語は脳内表象のネットワークの中に組み込まれる。これにより、りんごという音からなる単語の表象は、実際のりんごの表象と結合することで、果物としてカテゴリー化され、果物以外のカテゴリーと比べて、様々な果物とより強く連合する。言い過ぎかもしれないが、一旦カテゴリー化されると文法に当たるものは頭の中で過去の行為の記憶として成立している。「りんご(かき、ぶどう、梨、メロン)、食べる、美味しい」というわけだ。これを普遍文法の原型と呼んでもいいが、実際には単語が対応するものや行為にリンクした私たちの行動様式で、子供が最初に話すほとんどの言葉の並び(統語)はこの行動様式を反映していると思う。
このように、生物の寿命が伸び、複雑な内容の記憶が要求されればされるほど、mnemonicな連合相手が必要になる。この要求が、いつどのように言語という解決手段を見出したのか、それが次の問題になるが、今述べたのは個人の記憶の問題で、言語が発生する前も何らかのmnemonicな表象を使っていたと思う。しかしそれが言語になるためには、個人レベルのmnemonicな表象を他の脳とも共有できるようになる必要がある。そしてこれを考えるためには、もう一つの言語の機能、コミュニケーションについて考える必要がある。
ほとんどの生物が、何らかのレベルでコミュニケーションを行っている。それら全てを網羅して考えると複雑になるので、コミュニケーションを同種の高等動物同士が体を接しないで行う情報のやり取りに限定して考えると、1)子孫を残すためにオスメスが行う生殖行動に向けたコミュニケーション(生殖行為自体とは異なる、例えば発情期を知らせるサイン)、2)個体間の階層性を示すためのコミュニケーション、3)そして縄張りを相手に知らせるコミュニケーション、を動物にとって個体間のコミュニケーションが必要な3種類の状況と考えることができる。このときコミュニケーションが成立するためには、こちらが伝えたい内容について自分自身で理解しているとともに、同じ理解を相手も共有する仕組みが必要だが、この3種類のコミュニケーションでは、本能的感情が共有する情報の中心にある。メーティングや階層性についてのコミュニケーションは、生殖本能や、生存本能に基づく感情により動かされた行動だが、あらゆる個体が本能として等しく持っている感情(=情報)であるがゆえにコミュニケーションが図りやすい。
3つの中で、縄張りは直接対面しない個体間のコミュニケーションで、本能に基づく行為だが、様々なマークが縄張りの主張に使われる点で一層言語に近い。哺乳動物の場合、多くの動物の縄張りは、尿や糞など匂いの強いものを場所場所に残すことで主張される。これは、匂いが強い本能的感情を誘導するからで、このおかげで標識の意味を共有できる。クマのように、木に傷をつけて視覚的な標識を縄張りの主張に用いる種もあるが、実際には引っ掻いた場所に自分の皮膚や毛を擦り付けて残していることから、視覚的なシンボルに頼るわけではなく、印の付いた場所に匂いを付けて感情を惹起していることがわかる。遠吠えのような音で縄張りを主張する場合は、さらに言語に近いが、出し手と受け手が共有するのは、結局競争本能に基づく感情と言える。
一方多くの鳥類では匂いではなく、聴覚や視覚に訴える縄張りの主張が行われる。わが国で有名なのは、モズの高鳴きで、越冬場所を決めるまで鳴き声で縄張りを主張し続ける。他にも、極楽鳥の仲間のように美しい色を見せてメスを呼んだり、あるいはパッフィングと呼ばれる行動で相手を威嚇するなど多様で、これがバードウォッチャーを楽しませる(図2)。鳥の場合、いずれも感情を基盤とした本能的な行動であることがわかる。

図2:鳥の縄張り主張のための、モズの高鳴き(左)と、Victoria’s rifle birdのメーティング(左)。出典:Wikipedia
人間以外の哺乳動物と比べた時、鳥類には極めて複雑な音の並びを発することができる種類が多い。例えば、インコのモノマネからわかるように、鳥の中には様々な発声を学習することができ、その結果同じ種類でも場所によって鳴き方が異なるのを観察できる。しかし、その機能は生殖と縄張り、時に仲間に対する警告に限られており、共有する情報は本能的感情に限られる。しかし共有する情報が本能的感情だとしても、それに一つの表象を標識としてリンクさせる点で、形式としてはかなり言語に近い。(季刊生命誌70号では和多和宏さんによるキンカチョウの研究を紹介しています「小鳥がさえずるとき脳内では何が起こっている?」)
縄張り主張よりさらに高度なコミュニケーションが必要になるのは、社会生活を営み個体間で協力しあう時のコミュニケーションだろう。狼などの肉食動物や、チンパンジーの狩りなどが有名だが、この時のコミュニケーション手段はどれほど複雑なのだろうか?
例えばオオカミの群れが協力する狩りを考えてみよう(図3)。これまでの観察により、狼は多様な鳴き声を出せることがわかっている。このため協力して狩りをするとき、声を使って複雑なコミュニケーションを取っているのではと思ってしまうが、最近の論文では、狼が協力しておこなう狩りの途中ではそれほど複雑なコミュニケーションを必要としていないことが明らかにされている(MacNulty DR, Tallian A, Stahler DR, Smith DW (2014) Influence of Group Size on the Success of Wolves Hunting Bison. PLOS ONE 9(11): e112884. https://doi.org/10.1371/journal.pone.0112884)。すなわち、襲うという合図と、狩りに必要な自動的行動パターンを学習していることが重要で、狩りが一旦始まると個体間のコミュニケーションは必要ない。人間の争いでも修羅場になると、dog fightというようにコミュニケーション抜きで戦うのと同じだ。

図3:バイソンに立ち向かう狼の群れ:MacNulty DR, Tallian A, Stahler DR, Smith DW (2014) Influence of Group Size on the Success of Wolves Hunting Bison.
PLOS ONE 9(11): e112884. https://doi.org/10.1371/journal.pone.0112884の図1を転載。大きな群れでハンティングするときには、連携なく単独でのアタックでバイソンを倒すことが観察されている。
協力する狩りの例として有名なもう一つの例は、チンパンジーの集団によるヒヒの狩りだ。有名なジェーン・グドールの『In the shadow of man』にも、それまで毛づくろいをしていた集団が、急に立ち上がってヒヒを襲う行動を見せることが描かれているが、チンパンジーの狩りについての最も詳しい研究はBoeschらのコートジボアール・タイ国立公園での研究だろう(Boesch and Bosche, Hunting behavior of wild chimpanzees in the Tai’ National Park, American Journal of Physical Anthropology 78:547, 1989)。この論文によると、チンパンジーは実際の狩りより随分前から、胸をたたくドラミングやhootingと呼ばれる叫び声で狩りを始める合図を確認し、用意が出来たところで餌の声をキャッチすると、急に静かになって一定の距離を保って静かに獲物の方へと移動し、アタックすることが報告されている。さらに、タイ国立公園で観察されたグループでは、狩りの後獲物を奪い合うだけではなく、分け合うことも行われ、より高いレベルの協力関係が成立していると言える。さらには、追いかける役、行く手を遮る役などの分担も観察され、協力して狩りが行われており、獲物を食べる時の道具の使用も普通に観察される、知能的にも高度な集団と言える。しかし、狩り自体は、集団で狩りをするという意思の確認と、後は学習により身についた自発的な行動により行われていることから、役割分担といった複雑な関係を維持するために、コミュニケーションが存在しているのではないことがわかる。
以上のことから、集団の狩りは一見コミュニケーションが必要に思えるが、結局狼でも、チンパンジーでも、さらには私たちの先祖も、「ワー、オー」といった始まりの掛け声さえあればほとんどのことは済んでいたのではないだろうか。
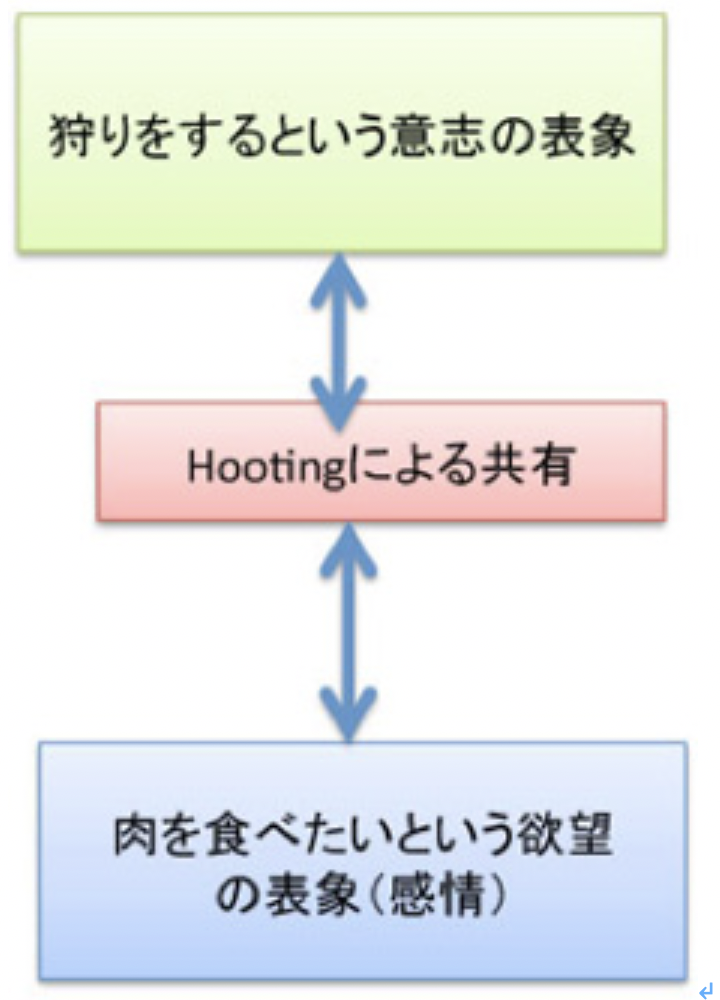
図4:感情と意志の関係を、hootingの音で他の個体と共有できる。感情の伝達はコミュニケーションの原点。
ただ、感情を基盤としたコミュニケーションを侮るわけにはいかない。ゆっくり鑑賞した絵画の詳細を思い浮かべることの難しさについて前回強調したが、感情を伝える最も高度な伝達手段の音楽では、一度聞いた曲が気に入れば、プロでなくともそのフレーズを正確に思い出すことができる。このことは、感情がもともとコミュニケーションを図りやすい表象で、例えば「これから狩りをするぞ」といった行動の意思統一も、肉を食べたいという強い感情が簡単なhooting音と組み合わさるだけで、比較的簡単に共有できる(図4)。
一方、得られた獲物を餌の分配する行動は、より高いコミュニケーション能力が必要とされる。この行動には、全体の、あるいは他の個体のために、自分は諦めるという利他性が要求される。そこで、次回は利他性とコミュニケーションについて考えるところから始める。
[ 西川 伸一 ]
私のような専門外の人間が言語について考え始めると、取り止めがつかなくなる。もともとこの連載は体系的に書いてきたわけではないため、言語のように難しい問題になると、問題が頭に浮かぶたびに脱線、脱線を繰り返してしまう。読者の皆さんを混乱させて申し訳ないと思っている。実際、自分で読み直しても、思いつきで右往左往、読んでいただいている皆さんを混乱させるだけで終わっている。ただ、とりとめなく文章を書いているうちに、それとはなしにホモサピエンスで起こった言語の始まりが自分なりにふっと頭に浮かび始めた。勝手に「言語のマイスタージンガー仮説」と名付けている。これを新しい年の冒頭に持っていきたいと準備しているので、それまでは、思いつくまま飛び回る私の思考を我慢してお付き合いいただきたい。
さて前回、狼やチンパンジーの狩りでは、相互に協力するための利他的コミュニケーションはほとんどなく、肉を食べたいという感情の共有と、狩りに必要な手続記憶に基づく自発的行動の学習、そして他のチンパンジーがどのような行動を取るかについての予測力で行動していることを述べた。このような狩りで情報として共有されているのはあくまでも感情や本能で、我々が考えるような感情以外の情報のやり取りが起こるためには、人類に特異的な質的に異なるレベルのコミュニケーションの進化が必要だった。
この人類(ホモ属)特有のコミュニケーション能力の背景について、類人猿と様々な発達段階の人間の児童を丹念に比べる研究で明らかにしようとしているのがライプツィヒのマックス・プランク進化人類学研究所のマイケル・トマセロ(Michael Tomasello)のグループだ。前回述べた複数の類人猿が共同で行う狩りの様式を、彼は人類の狩りと区別してexplosive(一旦スウィッチが入るとあとはコントロールできず爆発するだけ)な狩猟と名付け、類人猿には人間のコミュニケーションに見られる、相互作用による調整が全く欠けていることを述べている。
このサルから人類への進化過程で生まれた新しい個体間の協力関係が新しいコミュニケーションを発展させ、その結果言語の発生に至ったと考える研究者はTomaselloに限らず多い。ただ、Tomaselloはこのことを示すために、同じ課題を類人猿と様々な年齢の人間の子供に行わせて、人間にしかできない能力を科学的に特定する実験を数多く行ってきた。
例えば以前紹介した熊本のチンパンジー飼育施設で行われたTheory of Mind(他の個体も自分と同じように考えていることの理解)が類人猿にも存在することを示す論文は、熊本の施設だけで行われたのではなく、チンパンジー、ボノボ、オラウーンタンを用いてドイツ マックス・プランク進化人類学研究所のTomaselloの研究室でも同じ目的の実験が行われ、共同で発表されている。
この研究では、類人猿にもTheory of Mindが存在する、すなわち人類特有と思われていた能力が「猿にもある」ことを示している。確かにSF映画「猿の惑星」で人間と同じ知性を持つ猿が描かれていることからわかるように、ともすれば私たちの興味は「猿はどこまで人間と同じか」に向かう。しかし人類進化を考える上では、間違いなくTomaselloのように人間特有の能力であると検証できる性質、すなわち「猿にはできないこと」を一つ一つ明らかにすることのほうが重要だ。もともとできないことを証明するのは難しいため、このような実験結果は常に批判にさらされるし、普通すぐに「サルもできる」という論文が発表されることが多い。それでも頑なに両方を比べる実験を続け、サルと人間の子供の行動を知り尽くしたTomaselloの研究から学ぶことは多い。彼は多くの著書を出版しているが、Harvard University Pressから2014年に出版した『Natural History of Human Thinking』は、この他の個体との協力関係の変革から言語へと進む道がわかりやすく書かれており、人間特有の性質や能力とは何かに興味のある方にはオススメの本だ(邦訳がまだないのは残念だ)。

図1 Michael Tomaselloが2014年に出版した著作で、ここで説明している意図の共有を契機とした人間特有の能力の進化について書いている。
ではこれまでの研究から見えてきた人間特有の性質とは何か?またそれはどのように特定されたのか?誤解を恐れず単純化すると、Tomaselloを始めとする多くの研究者は、他の個体を助ける「利他性」を最も重要な人間特有の性質として挙げている。
もちろん他の個体を助けるかどうかだけで見れば、子供を育てる動物全てが利他性を持っていると言えるかもしれない。ただ、ここでいう「利他性」とはこのような本能的な利他性ではなく、本能的に持っている性質を超えて他を助ける利他性だ (例えば図2で施しと子育てを比較)。

図2:子育ても利他的だが、本能的と断じていいだろう。(写真はWikipediaより)
ではこのような高次元の利他性はどのように定義され、研究されているのか?わかりやすい例として、TomaselloのグループがPsychological Scienceに発表した「One for you, one for me: human’s unique turn-taking skills(一つを君に、もう一つは僕に:人間特有の順番性の能力)」という面白いタイトルのついた論文を取り上げてみよう(Melis et al, Psychological Science, 27:987, 2016 : http://journals.sagepub.com/doi/abs/10.1177/0956797616644070?url_ver=Z39.88-2003&rfr_id=ori:rid:crossref.org&rfr_dat=cr_pub%3dpubmed)
私たちは、協力しないと目的の物が得られない時、「最初に手に入れた物はまず君に、その次は僕に」と分配の順番を決めて協力することができる。しかしこの場合、一回の協力で手に入るのが一つだけだとすると、自分には何も手に入らないのに協力だけを行うという状況が生ずるのを認めなければならない。このためには、将来自分にも確実に回ってくる利益を想像して、一時的に我慢できる能力が必要になる。事実、3歳以前の幼児は、協力すれば欲しいものが手に入ることがわかっていても、自分に目的のものが回ってこないという状況を認めることができず、結局協力関係が成立しない。
この研究では、2本の紐を協力して引っ張れば、一個のボールを手に入れることができる2つの仕掛けを作り、片方の仕掛けからはボールが自分に、もう一方の仕掛けからはボールは相手に落ちるようにしておく。この使い方を理解させた後、2つの仕掛けを交互に操作して、順番にボールを手にするかどうかを、3.5歳児、5歳児、そしてチンパンジーで調べている。(同じ著者がProceeding of Royal Society Bに掲載したオープンアクセスの論文に、今回使われた仕掛けとよく似た図が掲載されているのでイメージを得るための参考にしていただきたいhttp://rspb.royalsocietypublishing.org/content/281/1796/20141973)。
図3: サルの協力関係を調べる課題。この図は、本文で説明している実験とは異なるが、紐を同時に引かせて協力させるという点では本文で説明しているPsychological Scienceの論文と共通してる。
結果は明瞭で、5歳児ペアの場合、実験開始直後からほぼ100%協力しあう戦略が成立する。すなわち、相手だけがボールを手に入れることを認め、交互に獲物が手に入るよう順番を決める戦略を立てる。ところが同じ戦略関係が成立する確率は、3.5歳児では62%に落ちる。この時の様子を観察すると、5歳児では積極的な方が声をかけて、順番にボールを手に入れようと指示しているのがわかる。一方、3.5歳児ではこの戦略が成立するまでに試行錯誤が必要で、結局うまく協力が成立しないペアもある。すなわち、この利他的能力の発達は3.5歳がちょうど境界上にあると言える。それでも、最終的には順番にボールを手にするための戦略を見つけるペアは何組か生まれる。これに反し、チンパンジーで同じ実験を行うと、偶然協力してボールを手に入れることは観察できても、順番にボールを手にするための安定した協力関係が成立することはない。
以上の実験から、Tomaselloたちは、将来の自分の利益を考えて他人とゴールを共有し協力しあう高次の利他性は人類特有の性質だと結論している。ただこの利他的性質が完全に自己の欲望を殺す道徳的行為の誕生と勘違いしてはならない。幼児はサルのexplosiveな協力形態と同じように、あくまでも自分の利益を追求しようとする強い利己的動機で行動しており、それでも協力が成立している点が重要だ。この実験で選んだペアも、自然に支配的な子どもと、それに従う従属的な子どもの区別が生まれて、支配的な方が行動を支持する。ただ階層性が存在しても、協力し合ったほうが一番得をすることが理解されており、協力を促す指示を出す。
この実験で観察された高次の利他性が人類特有の性質だとしても、なぜそれが言語発生までの長い道の契機になったのだろうか。極めて単純化して言ってしまうと、Tomaselloは人類(Homo)だけが、同じゴールを達成しようとする意図を共有する能力を発生させ、このおかげで新しいレベルのコミュニケーションが可能になり、また利他性もこの能力をきっかけに誕生したと考えている。前回述べたチンパンジーの集団的狩りで見られるexplosiveな協力でも、同じゴールを追いかけているように見えるが、実際には各個体は自分の欲求を満たすというゴールを追いかけている点で、意図を共有しているとは言えない。チンパンジーの狩りでは、各個体の脳内に表象されているゴールは利己的で個別の意図だ。強いて言えば、食べたいという強い感情を共通して持っているだけだ。従って、先に述べた実験からわかる、将来の利益のために他人だけが獲物を得るという状況を我慢できる利他性をチンパンジーは持ち合わせていないことになる。
私自身にとっても、この「意図の共有」能力が類人猿と、人類を分ける分岐点に存在するという考え方は納得できる。そこでTomaselloがBehavioral and Brain Scienceに発表した総説をもとに、「意図とは」「意図の共有とは何か」を考えてみよう。
例えば、近くで発見した鹿をなんとか殺して肉にありつきたいと考えている古代人を考えてみよう。彼の頭の中では、殺した鹿がゴールとして浮かんでいる。しかし、目の前の鹿は生きている。ゴールを達成するためには、その時取り得る幾つかの可能性を考え、鹿を殺すのに最も確率の高い方法を決断する必要がある。いろいろ考えた挙句、近くの石を投げて動きを止めて、あとは飛びかかって首を締めようと決断すると、ここで初めて行動する意図が生まれる。もちろんうまくいく場合も、うまくいかない場合もあるだろう。このような、ゴールを目指して現実を変化させことが、意図的行為になる。
こんな風に考えるプロセスは人間だけでなく、チンパンジーにも存在する。どちらもゴールを達成すべく意図的行為を繰り返し、その成否を記憶することで、鹿を手に入れるための方法を学習する。ただ、例えば石で鹿の動きが止まる確率が低いことがわかると、当然他の個体が棒を持って近づいた方がうまくいきそうだ。しかし一人では石を投げるだけしかできない。他の個体と協力するするしかない。
ではこのような協力を成立させるのに何が必要か?
図4 人間とチンパンジーの脳。構造は同じだが、前頭葉が人間では著しく発達しているのがわかる。この差は、人類が類人猿から別れた後、急速に進化してきたものだ。(出典:Wikipedia)
この意図の共有を新しいコミュニケーションの様式の発生と言っていいだろう。Tomaselloは意図の共有が発生する生後の発達についても詳しい研究を行っているが、詳細はBehavioral and Brain Scienceを読んでいただきたい。ただ、このような研究の背景には、Tomaselloに限らず、子供の発達を研究している研究者一般が持っている「系統発生は個体発生を繰り返す」という考えがあると思う。
そこで次回は、「意図の共有」を切り口に、コミュニケーション能力の個体発生と、系統発生の比較をしてみよう。
[ 西川 伸一 ]
自分自身で原著を確かめたわけではないが、「個体発生は系統発生を繰り返す」と反復説を唱えたのは、ドイツの生物学者エルンスト・ヘッケルだと習った。ヘッケルは19世紀ドイツを代表する発生学者で、名前に心当たりのない人も、おそらく彼が残した「Kunstformen der Nature」に描かれている様々な生物の精緻なスケッチは見たことがあるのではと思う(図1)。進化の過程で生まれた形態や機能(系統発生)は、個体発生過程を制限するため、個体発生では系統発生が短い時間に反復されることが多いという考え方だ。
ヘッケルも私たちの行動や精神にまでこの考えを拡大する意図は毛頭なかったと思うが、あえてこじつけて人間の行動の発達をヘッケルの反復説的に表現すると、「人間の行動の発達では、最初から人間らしさが現れるわけではなく、多くの動物と共通の本能的欲動に支配される口唇期を経て、行動支配の中心が前頭葉に移る中で、人間特有の性質である利他性などが芽生える」となるのではないだろうか。これを逆から言い直すと、利他性など人間特有の行動のルーツを探ると、私たちが動物の本能として理解している様々な行動が必ずその背景に存在し、個々の能力は、個体の発達過程で順番に現れることになる。

図1:エルンスト・ヘッケルとKunstform der Naturに描かれた図。(出典:Wikipedia)
私個人の感想でしかないが、脳機能が関わる限り、行動の発達が系統進化と同じ順番で現れるとする反復説がそのまま当てはまるかは疑問だと思う。というのも、脳はその時々の経験に応じてネットワークを書き換える能力を持っているからだ。確かに脳ネットワークの枠組みは進化過程で生まれてきたもので、個体発生でも脳幹に支配される行動から、より前頭葉が関わる行動へと発達していく。しかし、誕生後の行動や能力の発達過程では、脳内にある全てのネットワークが連合可能で、これに日々刻々積み重なる外界の記憶も必要に応じて連合される。このため、系統発生過程で形成された様々な本能の回路も、新しい経験や、認識や価値の回路と自由に結合する可能性が常に存在する。例えば、口唇期をガイドするのは食欲本能が中心だと思うが、母親の匂いや、声を同時に聞くことで、本来ずっと後で発生してくる生殖本能が連合される可能性すらある。もちろん、新しい経験が思いもかけない連合を誘導する可能性も十分ある。
系統発生の結果として始まる最初の脳ネットワークの構造の個人差は大人と比べるとはるかに少ないと思うが、その後の経験は偶発的で多様だ。従って、ネットワークの書き換えも、経験の個別性の結果大きな多様性が生じてしまう。系統発生で獲得された本能のネットワークはどんなに共通でも、様々なネットワークと自由に連合させ、新しいネットワークへと再統合することができる。まさにこれが、人間の個性の源で、例えば生殖行動から子孫を作るという目的を切り離してしまうことも、その結果として性同一性障害が生まれることも、全て脳内でのニューラルネットワーク同士を自由に連合させられる特性に起因している。この意味で、行動の個体発生は、必ずしも行動の系統発生を正確に繰り返す必要はない。
しかし、脳のネットワークを自由に書き換えられるからといって、系統発生で獲得された本能から完全に解放されるかと言うと、病的なケースを除いて、系統発生で生まれた行動は、発達中のネットワークに統合され必ずどこかに潜んでいる。例えば、言語発生にはサルにはない人間特有のコミュニケーション能力が必要なこと、このコミュニケーション能力は人間特有の高いレベルの利他性がきっかけになっていることを説明した。では、この全く新しい人間特有の能力は、サルや他の哺乳動物に見られる本能から完全に独立しているのだろうか?
答えは、残念ながらNOだ。人間特異的な高次の能力や行動には、それに対応するより原始的な能力が必ず存在している。この意味で、人間特有の全く新しい能力の理解のためには、系統発生と個体発生を対応させ、人間特有の全く新しい能力(これは系統発生では辿れない)が、他の動物にも共通に存在する能力とどう関連して発達してくるのか研究する必要がある。
利他性を例に、さらに人間特有の能力と、他の動物に見られる能力との関係を見てみよう。利他性というからには、当然自分と他の個体を区別することが必要になる。
自分自身を認識できるか(self awareness)調べるために行われるのが、鏡に映った自分の体を自分と認識できるかについての実験で、例えば背中にこっそり印をつけておいて、鏡を見せた時にその印に気づいて自分の背中に注意を向けるかどうか実験する。人間の場合、2歳時以降からこの能力が認められ、さらに4歳児ではビデオで見た自分の姿も認識できる。一方、チンパンジーなどの旧世界ザルでもこの能力が認められるが、ビデオを通して見せた場合は、霊長類も自分とは認識できないようだ。また、より原始的な新世界ザルでは鏡に映った姿を決して自分と認識することはない。
自他が区別できるようになった後は、他の個体の行動を理解できるかが重要になるが、これについて最も有名な研究がミラーニューロンの発見だろう。このニューロンは、霊長類の餌をとる際の手のニューロンの活動を記録する実験を行う最中に、実験を行う研究者が餌のバナナをとったのを見たとき、必ず活動する神経を見つけたことに始まる。すなわち、相手の行動をあたかも自分の行動のように反応するミラーニューロンの発見は、20世紀の神経生物学の最も重要な発見の一つと考えられている。人間の場合、霊長類のミラーニューロンに対応するニューロンを特定することは難しい。ただ、相手の行動を目で追いかける行動は1歳児までに見られることから、同じようなニューロンは間違いなく存在すると考えていいだろう。
自己と他を区別できるようになると、次は他の個体も自分と同じように考えているという認識(Theory of Mind)能力の発達が重要になる。これについては既に詳しく述べたが(http://www.brh.co.jp/communication/shinka/2017/post_000016.html)、最初は人間特有の性質と考えられてきたが、その後の研究で霊長類にもその能力が備わっていることが証明された。また、人間でも社会性に問題がある自閉症の子供たちにはこの能力が欠けていることも明らかになっている。
上に述べた他の個体の行動を理解する、いわば理性的能力の駆動力として欠かせない情動が、相手の感情を共有する共感能力だ。この共感こそが、他の個体とのコミュニケーションを図ろうとする原動力になり、この能力が低下するとAlexithymia(無感情症)に陥る。ただ、共感能力は人間以外の動物にも存在する。例えば、相手の痛みを共に感じる感情は、霊長類は言うに及ばず多くの哺乳動物で観察することができる。例えばマウスでも、痛みを訴える仲間のところにより頻回に寄り添う行動を示すことが観察されている。このことから、一定の集団で暮らす哺乳動物では、他の個体とのコミュニケーションを求める情動が様々な形で早くから進化していたことがわかる。
それもそのはずで、妊娠、出産を経て一定期間子供を育てなければ子孫が残せない哺乳動物の子育て(ケアリング:caring)には、子供への共感が必須だ。ほとんどの動物で、ケアリングは本能的な行動で、子供の発する声などの刺激を受けて活性化される視床・帯状皮質を中心とする辺縁系の進化により可能になっている。人間の子供への愛情も例外ではなく、同じ辺縁系からの情動なしには維持できない。しかし、人間の場合この辺縁系の本能のネットワークを様々な前頭葉皮質領域が連合することで本能とは別の行動が可能になっている。例えば、自分の子供以外をケアリングしたり、あるいは人間以外の動物の子供のケアリングもその例だろう。
ケアリングに関わる共感は、さらに進むとオスとメスの間の性行動や、つがいの形成(Pair bonding)にも見られるようになる。もちろんこの共感も、子孫を残すという目的を最も効率よく果たすよう進化した脳幹の回路に支配されており、本能の回路だ。実際、広く動物を見渡しても、子孫を残すという目的以外で生殖行動を起こすようになったのは人間だけではないだろうか。
さて人間のPair bondingの特徴は、一夫一婦(monogamous:もちろん例外もあるが)だろう。一般的に、子孫を残す目的の生殖行動は、類人猿を含む大半の哺乳動物で、強いオスだけが生殖本能を満たして子孫をのこす一夫多妻であることが多い。人間に近いチンパンジーもこのスタイルをとっている。攻撃性がなく、時に利他性すら示すとして、道徳の起源を探ろうと研究されているボノボですら、一夫一婦制の家族を作ることはない(極端に言うと相手を選ばない乱交型と言えるかもしれない)。しかし、一夫一婦制をとるのは人間だけではない。5%弱の哺乳動物では生涯一夫一婦のつがいで暮らすことがわかっている。すなわちこれらの動物では、新たな生殖行動の様式として一夫一婦制を進化させてきたと考えられる。残念ながらなぜこのような一夫一婦様式のpair bondingが進化したのか答えることができないが、このpair bondingにはそれ以外の動物にはない仕組みが存在することはわかっている。

図2:プレーリーハタネズミ。(出典:Wikipedia)
一夫一婦型の動物として、最もよく研究されているのが、Prairie Vole(プレーリーハタネズミ)だ。社会性を誘導するホルモンとして有名なオキシトシンをメスの脳に注射すると、出会ったオスとの絆が強まることが知られている。一方、オスのメスに対する絆を強めるためには、オキシトシンの代わりにバソプレシンを脳内に注射する必要があることがわかっている。逆に、オキシトシンやバソプレシンの作用を阻害する分子を脳内に注射すると、pair bondingを阻害することができる。すなわち、ハタネズミは進化の過程で、オキシトシン、バソプレシン刺激に反応する神経ネットワークを、生殖本能のネットワークと連合させることで一夫一婦システムを進化させたことがわかる。
全く同じとは言えないものの、人間の一夫一婦型ペアリングにも同じ原理が残っていることが知られている。少し浮世離れした面白い研究で、初めて読んだ時私も「大笑いした(?)」3題話になっているので詳しく紹介しよう(図3)。

図3 米国アカデミー紀要に掲載されたPare bondingに関わる3編の論文。
発端は2008年1月発行の米国アカデミー紀要に掲載されたフロリダ大学の論文だった。論文の内容は驚くべきもので、実験室で飼っているハタネズミのオスの中に、他のメスと浮気をする個体がいるので、そのバソプレシン受容体(V1aR)の脳内での発現場所を調べると、一途に添い遂げるネズミと比ベて大きな差が見られることが報告されていた。
これでも十分面白いのだが、同じ年9月にスウェーデンカロリンスカ大学から、V1aRの遺伝子多型と男の浮気心についての研究が米国アカデミー紀要に発表された。この研究では、V1aRの一塩基多型(SNP)を調べ、特定のSNPを持っている男性は離婚を含む結婚の危機を経験している確率が、そのSNPを持たない男性と比べて2倍高いことを報告している。さらに、結婚ではなく同棲している率もこのSNPを持つ男性の方が2倍多い。この結果は、人間の夫婦形態が教育、文化等の影響を受けて複雑になってはいても、本能的なところでは、ハタネズミと同じ情動が働いていることを示唆している。
これらの結果は、男性の特定の女性へのpair bondingに、ハタネズミと同じバソプレシンが関わることを示しているが、人間の進化の過程で、オスはバソプレシン、メスはオキシトシンという差を失いつつあり、男性のpair bondingにもオキシトシンが効果を持つことがドイツ・ボン大学から。2013年11月にやはり米国アカデミー紀要に発表された。実に楽しい実験で、結婚前の恋愛進行中の男性を集め、恋人の写真、及び無関係の魅力ある女性の写真に対する反応を機能的MRIで調べている。写真を見せる前にオキシトシンスプレーを鼻に投与する群と、偽薬を投与する群に分けて、ステディーな恋人の写真を見た時に興奮度を高める効果がオキシトシンにあるかどうかを調べている。専門家でないので、どの程度MRIのデータを信用していいのかはわからない。ただ結果は予想通りで、オキシトシンを投与されると、今つき合っている恋人の方により強く興奮するが、偽薬だと恋人の写真と、知らない女性の写真を見たときの反応に大きな差が無くなると言う結果だ。
言語誕生を人間の社会性と一夫一婦の夫婦形態が促したと考える研究者は少なくないが、動物共通の共感本能から発展したpair bondingの進化を探るだけでも、本能に関わる脳ネットワークを基礎に、それをさらに支配するためのネットワーク間の連合が進化していることがお分かり頂けたと思う。脳というシステムの性質上、系統発生で進化した能力がそのまま個体発生で繰り返すことはないとしても、人間特有の能力のルーツを辿れば、脳の進化過程で拡大してきた本能の進化があることは間違いない。この意味で、人間の発達過程で起こる現象を整理して、系統発生的ルーツを考えることは重要だ。
言葉の発生に戻ると、育児書では言葉の発達は幼児語が始まる1歳前後から始まるが、もっぱら泣いているだけに思える3か月ぐらいからすでに、少しずつではあるが周りで起こっていることが理解され始め、泣くという行為を通して、コミュニケーションを図ろうとしている。その後。「バババ」と言った赤ちゃん言葉(babbling)を経て、ママ、パパのような意味のある言葉が発展していく。この時、お母さんから見るとどうしても言葉を話すということに注意が集中してしまうが、実際には周りの出来事についての理解、様々な方法でのコミュニケーションの試み、社会(人間関係)の認識などが脳発達とともに進行している。これまで読んだ論文の印象から言うと、この過程の詳しい記述は始まったばかりではないかと思う。その意味で、今後人間の個体発生の記録の蓄積は、まだまだ言語誕生の研究に欠かせない。特に最近の赤ちゃんは、私たちの時代とは全く異なる家庭環境に置かれている。この新しい状況が、系統発生で獲得された能力にどう影響していくのか、フロイド、ユング、ピアジェの時代の個体発生過程の記述を比べることで、予想もしない発見があるかもしれない。
次回は、これまでのまとめとして、私が言語発生の過程についてどう想像しているのか「言語発生のマイスタージンガーモデル」というタイトルでまとめてみたい。
[ 西川 伸一 ]
これまで言語誕生について、断片的に書き留めてきたことの中間まとめを2回に分けてお届けする。脈絡なく書き留めてきた文章になんらかの筋を汲み取ってもらえればありがたい。一種の書き下ろし原稿になっているので、この文章だけで十分理解してもらえると思う。またこの原稿を通して、言語を考える時に参考になる本を紹介する。
言語誕生を促す2つの契機
互盛央さんの「言語起源論の系譜」(図1)を読むと、人間は言語を話す自分に気がついてからすぐ、自分の使っている言語はどのように生まれたのか考え続けてきたようで、この問題は少なくとも2500年の歴史がある。しかし、人間の脳との関わりで議論が始まったのはやはり20世紀で、中でも私たちの言語能力は決して学習するものではなく、生まれつき備わっているものだとしたチョムスキーのUniversal Grammar仮説は影響力が大きく、言語の起源をめぐる議論の中心になった。

図1互盛央さんの『言語起源論の系譜』(講談社)
言語起源論の歴史を知るには面白い本で、深い知識に裏付けられている。
チョムスキーは、言語発生がコミュニケーションの延長線上にないことをことさら強調することで、universal grammarを前面に出して言語の発生が統語能力の発生であるとする論陣を張った。これに対し、言語発生には、人類特有の新しい社会関係を支えるコミュニケーション能力が必要だとする陣営は黙ってはいなかった。現状はというと、(個人的印象だが)言語誕生の背景に人類特有のコミュニケーション能力の進化があると考える人の方が今では多くなっているように思う。
この論争についてここで詳しく解説する余裕もないし、またどちらを支持するのかという議論をするつもりはない。代わりにこの原稿では、普遍文法もコミュニケーションの能力も、言語誕生に必須の独立したモデュールとして考える。すなわち、チョムスキーの普遍文法が提起した統語の問題を、行動についての表象能力の問題、ボキャブラリーの問題を作業記憶過程での連合の問題、そして言語の誕生に関わるコミュニケーションの問題を、自分を含む社会のゴールの表象の問題として捉え、これらが交わるところに言語が発生したという観点から考えてみたい。
比較進化学として言語誕生を考える
言語は人間特有の能力で、地球史におけるそのインパクトは、地球上にこれほど多様な生物をもたらせた進化を可能にした生命最初の情報DNAにも匹敵する。言語を持つ人類ののみが文明を発展させ、他の生物を圧倒する繁栄を遂げた。その結果は地球の環境も変化させる力がある。例えば、もともと地球上にO2と表現される自由酸素はほとんどなかった。しかし、生物が誕生し、その後光合成を行う生物が進化すると(おおよそ35億年)地球上でも酸素が作られるようになり、生物の繁栄とともに10億年前から急速に濃度が上昇した。この変化は全て、進化を可能にしたDNA情報の誕生の結果だ。
同じように、現在地球温暖化の元凶と言われる2酸化炭素の蓄積をもたらしている工業化の問題は、元をたどると言語という全く新しい情報媒体誕生まで遡ることができるだろう。言語誕生を5万年から10万年と考えると、これほど短い期間に人類を繁栄させ、地球の生物相や大気成分までを大きく変化させる力を言語は持っていたことになる。事実、千年前は人間、家畜、ペットの哺乳動物に占める割合は1%に満たないと推定されているが、現在は何と98%に及んでいる。これは決して人間の筋力が他の動物に比べて強いからではなく、人間だけが言語を獲得した結果に他ならない。
このように人類だけが地球上で言葉を使い、文明を発達させ、地球の大気すら変化させた。では言語はどのように現れたのか?この過程を「創発」という言葉で終わらせるのは思考の停止だ。実際には見ることができない過去に起こった言語誕生を理解するために、現存の人間特有の能力と、他の動物、特に我々に最も近い霊長類の能力を比べ、それぞれの違いを理解した上で、各能力と言語の関わりを探る研究が続けられている。
言語誕生に関わる変化のうち一番わかりやすいのが、喉頭の解剖学的構造だ。口腔から離れた下の方に喉頭がある人間と比べると、霊長類の喉頭は口腔・鼻腔直下にあるため、声帯で発生させた音を口腔内で操作することができない。結果、私たちのように多様な母音や子音を発生することはサルには不可能だ。サルが発生するほとんどの音は鼻で増幅する鼻音として発声される。このため言語に必要な複雑な音を発生することは霊長類ではできない。
ただ、発声に必要な解剖学的特徴は言語に必要なモデュールのほんの一部でしかない。もっと重要なのは、記憶とコミュニケーション能力に起こった変化で、この条件が揃わないと、オオムと同じことになる。手短に、人間特有のコミュニケーションと記憶能力について見ておこう。
目的と意図を共有する新しいコミュニケーション様式の誕生
映画「猿の惑星」でサルも進化すれば人間と同じ能力が獲得できると描かれているように、私たちの興味はともすると「サルはがどこまで人間に近づけるか」、すなわち「サルでもできる」ことに向きがちだ。例えば、膨大な努力を払って、チンパンジーやボノボに言葉を教える研究がその例だ。これまでサルが100語以上の単語を正確に区別でき、覚えた単語を使って意思表示も可能であることなど示されている。しかし、言語誕生の条件を考える上で本当に重要なのは、間違いなく人間にはできてサルにはできないことを明らかにすることだ。実際には、できないことを証明するのは難しく、また一つの仮説の検証に、時間をかけてサルを訓練する必要があり、手間のかかる大変な実験だが、この努力のおかげで「サルにはできない」幾つかの人間特有の能力が特定されてきた。中でも、ライプツィヒのマックス・プランク進化人類学研究所(Max-Planck Institute of Evolutional Anthropology)のマイケル・トマセロ(Michael Tomasello)は、同じ課題をチンパンジーなどの類人猿と、様々な年齢の人間の児童に行わせて、人間特有の能力とは何か、またそれがいつの段階で獲得されるのかを丹念に調べている。彼が2014年に出版した『Natural History of Human Thinking』はこの分野に興味のある方には是非読んでいただきたい著書だ。

図2:Tomaselloが2014年に出版した人間の思考についての著作『Natural History of Human Thinking』(Harvard University Press)。多くの実験事実の裏付けられており、アカデミックな読み物。
Tomaselloをはじめ、この分野の多くの研究者が一致しているのは、人間特有のコミュニケーション様式を生み出した社会性が「一種の利他性」だという点だ。しかし、利他性なら母親が子供の面倒をみるcaringや、オスとメスがつがいを作って(pair bonding)暮らしている動物にも存在するはずだ?
たしかに、人間の持つ利他性の究極の起源にはcaringと同じ情動の仕組みがある。たとえば一見利益の反する行為も促すことができる「ご褒美回路」として知られている仕組みは、人間の利他性にも働いていることはまちがいない。しかしサルと人間を比べると、人間の協力関係だけに、「他の個体とゴールと意図を共有する」という特徴が見られ、これが動物の利他的行動には欠落していることがわかる。
Tomaselloの実験の一つを紹介しよう。お菓子を手に入れるのに協力が必要だが、得られるお菓子は一回に一人分しか出てこない機械を挟んで座った2人の子供、あるいは2匹のチンパンジーの行動を観察し、お菓子を順番に手にするために、相手が先にお菓子を手にするのを我慢できるか調べた実験を行うと、人間の子供は(3.5歳齢以上)協力して「今は僕、次は君」というように順番にお菓子を分けるのに、チンパンジーでは偶然に協力関係が成立することはあっても、自分が我慢を強いられる順番があることを受け入れられず持続的な協力が成立しないことを示している(Melis et al, Psychological Science, vol 27, issue 7, 987-996, 2016)。
集団行動という点で見れば、チンパンジーなどの霊長類も肉を求めて複数の個体が協力して小さなサルの狩りを行うことはよく知られている。しかし、一見協力して行われている狩もよく観察すると、各個体は自分が獲物にありつくという目的だけを考えており、獲物を得て分け合うという共通の目的を共有することはできない。
共通のゴールを共有する協力関係には、個体同士で指示をし合うコミュニケーション、すなわち相手の意図が理解でき、自分の意図を相手に理解させることが必要になる。このような指示関係が最初は自然発生的に成立しても、同じことを繰り返すためには、上記の論文のタイトルにあるように「今度は君、次は僕」といった取り決めを両方が確認する必要がある。先に述べたTomaselloの論文でも、協力し合う3.5歳児は、指差し(ポインティング)を用いてコミュニケーションを図っていることが記載されている。このように「君と僕」といった2人の間の比較的簡単に見えるコミュニケーションでも、人間だけにしか、しかも3.5歳児になるまで観察することができない。もっと多くの個体同士がゴールを共有して協力するとなると、さらに高次のコミュニケーションが必要になることだろう。
ではこのような意図の共有はどのように発生してきたのだろう?もちろん正解があるわけではないが、多くの研究者は共同生活するグループの人数が増え、社会構成が複雑になったことが大きな原因だとかと考えている。この根拠として最もよく引用される研究がロビン・ダンバー(Robin Ian MacDonald Dunbar)らが1992年に発表した論文だ(Dunbar RIM, Neocortex size as a constraint on group size in primates, Journal of Human Evolution 20, 469-493, 1992)。
この論文の要点は、霊長類から現代人までの新皮質のサイズと共同生活での集団のサイズをプロットすると両者が正比例することだ。すなわち、集団構造の複雑性と、脳のサイズが正比例することを示している。アフリカでの最初の人類誕生はおおよそ400万年前だが、脳の新皮質の急速な増大がみとめられるのは200万年後、Homo.Ergasserが誕生してからで、彼らのグループの人数も、それまでの50人以下から、100人近くに倍増していると考えられており、Dunbarの仮説を支持している(図3にDunbarの最近の著書1冊を紹介しておく)。

図3 Dunbarが2014年に出版した『Human Evolution』(Pelican)。何が我々人間の進化を後押ししたのか、言語誕生に至るまでわかりやすく書かれている。
最初の原人がアフリカに誕生したあと200万年間、人類のハンティングは、死体や食べ残しの骨をかすめるハイエナのような狩りだったと考えられているが、H. Ergasserになると人間より大きな獲物を求める狩りが始まった。石器の進歩は10万年前まではゆっくりだが、それでもH.Ergasserの石器には動物を殺すための進歩が見られる。いずれにせよ、100人程度の共同生活が可能になるためには、野生のチンパンジーの狩りとは違って、獲物を分け合う集団生活が行われていたと考えられ、ゴールや意図を共有して協力する社会が形成されていた。
重要なことは、この複雑化した協力関係や利他性の背景には、必ず新しい表象と統合システムが脳内で形成されているという点だ。このシステムの要件として、

図4 Mithenが2005年に出版した歌うネアンデルタール『The Singing Neanderthals』(Harvard University Press)。言語誕生に音楽が果たした役割を示す面白い考え。
しかし、意図を共有したコミュニケーションを可能にする脳構造のおかげで、集団的な狩りの成功率は上昇し、その結果タンパク質の豊富な肉の消費量は上昇し、脳の拡大は更に加速したことだろう。このように、5−10万年前まで、集団で獲物を追いかけるHunter Gathererが目的を共有し分業により獲物をしとめる暮らしには、私たちが考えるような複雑な言語がなくても、ジェスチャーやHmmmmmで十分済ますことができたと思われるが、おそらく脳自体は大きくなり、より言語の誕生を支える可能性が高まったと思われる。
コミュニケーションを求める力動は言語発生の必須条件
では、ゴールを共有する社会性を持つことが言語発達にどのような影響を持つのだろう?進化の過程でこれを検証することは、現在はまだ難しい。しかし、人間の言語発達過程にこの社会性が重要な役割を持つことは、自己と社会との表象が変化する自閉症スペクトラム (ASD)やウイリアムズ症候群(WS)の解析を通して認められているので、短く紹介しておく。
ASDは、社会性の低下、言語発達障害、そして反復行動を主要な症状として示す発達障害を指し、症状は同じでもその原因は極めて多様であると考えられており、そのため自閉症という病名ではなく、自閉症スペクトラムと称す流ことになっている。原因はともかく、知能の発達が正常でも、社会性の低下と、言語の発達遅延が必ず合併していることから、他人とのコミュニケーションを求める力動が言語発達に必要な条件であることを示す例として考えられてきた。事実、ASDは乳児期から能動的に体験を求め外界へ働きかける行動が見られないことがわかっている。正常児では生後3−4か月までに相手の顔をじっと見つめ、動くものを目で追いかけ、音がする方向に顔を向ける動作が見られる。ところがこのような反応がASDではしばしば欠けている。このため米国疾病予防管理センターは、「もしあなたの子供が、音に反応せず、動くものを目で追いかけず、笑わず、指をしゃぶらない」なら、すぐに専門医に相談するよう勧告している。おそらく胎児発生の過程で形成される脳の回路が普通とは違っていたため、外に経験を求める力動が低下して、コミュニケーションを求める欲求が減ったため、結果としてコミュニケーションの媒体である言語発達を求めないのだろうと考えられる。
このような解釈は、都合の良い現象を集めただけの、思弁的な想像だと思われても仕方ないが、もう一つの例ウイリアムズ症候群について知ると、まんざら間違っていないように思えてくる。
ウイリアムズ症候群は1961年外科医により「大動脈弁上部狭窄症と特徴的精神症状を示す」一群の患者の存在として記載された。その後の研究で、7番染色体上の大きな欠損によりその領域にある20種類の遺伝子が片方の染色体ごっそり抜け落ちてしまっていることがわかった遺伝疾患だ。もちろん病気の詳しい解説をするつもりはない。重要なのは、この患者さんたちが自閉症とは逆に「相手の目をじっと見つめる、愛すべき人懐っこさを持つチャーミングな性格」を持つことと、知能の発達が障害されIQは60程度であるにもかかわらず、言葉を話す高い能力を持っている点だ。実際には、知能発達の遅れのため言葉を話し始める時期は遅れるのだが、一旦言葉を話し始めると多弁で同年齢の通常児を凌駕する。すなわち、社会性と言語能力の関係という観点から見た時、ウイリアムズ症候群の患者さんには自閉症とまるで反対の症状が併存している。このことは、「愛すべき人懐っこさと称される高い社会性」が、知能発達遅延による言語発達への影響を克服できるだけの力があることを物語っている。
これらの例は、個体の発達から見ても、コミュニケーションを求める力動、すなわち社会性が言語発達の条件になっていることを示している。この意味で、言語のコミュニケーション機能を無視する考えには賛成できない。
※「言語誕生のマイスタージンガーモデル(後編)」は2018年2月1日に公開します。
[ 西川 伸一 ]
音節と記憶の連合(Lexicon:語彙は、連合させて記憶を高めるメモとして
始まった。)
話題をガラッと変えて、次は記憶について考えてみよう。私たちには瞬間ごとに、膨大な量の情報が感覚を通して入ってくる。これら全てを記憶することは不可能だし、ほとんどは意識にすら上ることなく通り過ぎる。意識されるのは、各人が経験で形成してきた独自の基準とその時の脳の状態に照らして選択された表象だけで、この時の選択を行う基準を、「自己」と呼んでいいと思うが、自己の表象自体も刻々変化する脳内ネットワークだ。
感覚を通して入ってくる情報
の中から特定の表象を選択して記憶しているとは言っても、物事を正確に覚えているのは難しい。我々の感覚の7割は目を通して入ってくるが、例えば初めての美術館を巡って外へ出た瞬間、目の前にもう一度イメージを思い浮かべられる絵は何枚あるだろうか。実際、覚えられないのが当然で、絵を見るとき私たちが画像をどのように脳内に表象しているかを考えれば、覚えていることの方が奇跡だ。実際には、絵の様々な場所に視線を走らせて、そうして得られた各部分の表象を、形、色など様々な要素に分解し、それを脳内の異なる部分に維持した上で、もう一度脳の中で表象へと統合し直す。すなわち、絵を見ている瞬間として認識している体験も、実際には細切れになった様々な表象を脳内各所に維持し、それを再統合したものをある瞬間に見た画像だと認識しているだけだ。この過程は、作業記憶と呼ばれているが、この複雑な過程を何回も繰り返すのが絵画の鑑賞になる。とすると、情報量が多すぎてなかなか記憶に残らないのも当然だ。しかし見た瞬間、なぜか深い感動を覚える絵は間違いなく記憶している。これは複雑な視覚の表象でも、感情という単純な表象と連合させると覚えやすいからだ。もちろん、連合相手は感情だけではない。何か単純なシンボルとうまく連合させれば記憶が容易になることは多くの人が経験していると思う。美術館で写真を撮ったという記憶があると、その絵を覚えている確率が高まるという研究があるが、これも写真を撮るという簡単な表象と連合させているからだ。
匂い、音、驚き、恐怖の経験など、感情的表象は記憶を容易にする連合相手の定番として実際使われているが、短い音節も記憶に残りやすく、連合相手として優れていのではないだろうか。私たちはすでに言語を持っており、言語に似た音節を個人的な記憶を高めるための個人的メモ代わりだけに利用している原始的状況を想像できない。しかし、私たちの記憶に、実際の事物の表象と音節を持ったシンボルとしての単語の表象の連合は大きな役割を果たしており(固有名詞と顔の表象を考えてみてほしい。言語的連合があると覚えることができる)、個人的に、特定の音節を連合させて
記憶を高めるために使うことは十分あり得るのではないかと思っている。言語が誕生する前は、この音節と事物の表象との連合は偶然結びついただけで、シンボルとして表象に固定されているわけではないし、ましてや他の個体や社会と共有しているわけではない。しかし私たちの脳内には、語彙とは全く別に様々な事物についての表象が階層的に形成されている。例えば、食べられるもの、食べられないもの、甘いもの、しょっぱいもの、といったカテゴリーだ。この表象に、音節がいったん連合されると、当然その音節は、頭の中に形成されている複雑な表象のネットワークに統合される。
もう少し具体的に見てみよう。言葉を持たない原人でも、獲物の鹿が見つかった時には思わず声が出る。この声になって出た「ディア」といった音と鹿と周りの景色を連合させたとしよう。すると、この「ディア」という音は、鹿を見つけた経験の表象と個別に連合されるが、これにより原人の頭の中にあるほとんどの表象のネットワークに組み込まれたことを意味する。例えば「ディア」という音節は鹿とその周りの景色というだけでなく、ライオンとは違うこと、食べれば美味しい、あるいはもっと大きなカテゴリー、4つ足の動物などの表象とも関連することになる。
連合による明瞭な記憶は自己自身の表象の確立に始まった。

図1 昨年出版された、Murray, Wise, Grahamの3人の共著による記憶の進化についての著書『The evolution of memory system』。進化という観点で記憶を捉えた本としては初めてではないだろうか。
上記のように、物事を明瞭に覚えるためには、関係のない表象を連合させられる能力が必要になる。異論もあると思うが、このような能力が人類だけに発達したと考える研究者は多い。中でもエリザベス・マレー(Elisabeth Murray)等によって最近出版された『The evolution of memory system』では、明瞭なexplicit memoryと呼ぶ明瞭な記憶を形成する能力は人類にしかなく、類人猿を含む動物では発達していないと主張されている(図1)。というのも、explicit memory形成には、新しい連合とそれを統合する脳の仕組みが必要で、具体的には考え行動している自己を身体認識とは切り離して表象し直し、エピソード記憶形成過程で、一旦分解された感覚からの表象を、もう一度統合する基準にすることが、人類だけで可能になったからだと主張している。一種の、主観的意識のようなものだと思うが、説得力のある主張だ。
動物にexplicit memory(顕在記憶)やsemantic memory(意味記憶)があるのかないのか、全く判断できないが、それでもこの本の指摘は言語を考えるためには重要だと思う。というのも、自分が発生した音節を、全く無関係のエピソード記憶と連合させ記憶を鮮明にする、音節をメモ代わりに使う能力は、explicit memoryを可能にする能力と完全に重なるからだ。この本では、言語であれ、新しい文明であれ、すべては人類だけが獲得した、explicit memory形成能力に起因するとまで提案している。確かに、homo sapiensが生活していた洞窟に残された動物の絵は、明瞭な記憶なしに描くことはできない。
さらに、カテゴリー化の能力や、全く無関係な表象を連合させる(例えば母親(mother)、と自然(mother nature))semantic memoryも内的自己の表象を、様々な表象を連合させる基準に据えることで可能になったと考えられる。要するに、偶然であっても、自分を中心に全く関係のない表象と表象を連合させる能力は言語発声にとって必須の条件になる。
しかし特定の音節をいくら頭の中の様々な表象と階層的に連合させられるとしても、その過程を他の個体と共有できているわけではない。言語発生には、この音節と事物の表象の関係を複数の個体が共有する必要がある。この問題については最後に議論する。
行動の表象が文法を決める
言語発生前の話をしていることを再確認しておく。したがって、ここで議論している言語は、たかだか語彙が100にも満たない原始的なものだと考えておいてほしい。この状況を念頭に次に文法について考えてみよう。
3歳前後に、最初に自発的言葉を話すようになっても、使う単語は3−4語程度だが、例えば「水・飲みたい」とか「表・出たい」、あるいは名詞を並べて「ママ・今・部屋」などだ。重要なことは、誰が聞いても意味が十分くみとれる言葉を話している点だ。これをもって、普遍文法構造が脳内に形成されていると結論しても悪くはないが、このような幼児の言葉の統語を、私たちが何かをするときかならず頭の中に形成する行動の表象に従って単語が並べられている結果と考える可能性はないだろうか?私たちの行動は、時間的にも、空間的にも様々な制約を受けている。この行動に対する制約は、一つは意識には登らない手続き記憶とし、もう一つは意識された行動の記憶として脳内に表象されているはずで、原始的な言語ではこの表象がそのまま統語のルールとして使われている可能性がある。
実際、運動と文法の間に関連があると考えられる現象は存在する。この関係が最も明確に見えるのが、運動の中枢、小脳の障害により起こる失語だ。小脳性の失語は、有名なブローカ失語やウェルニッケ失語とは症状が異なり、Agrammatism(失文法)と呼ばれる文法の異常が見られることが多い。このことは、小脳内に統合されている行動の表象が文法としての機能を果たしている可能性を強く示唆している。
また、大脳皮質の失語に失行症が併発することもある。失行症とは、自分の行動についての表象が障害される疾患だが、この結果も行動の表象と言語が強く連合していることを示している。このように、原始的な言語に見られる普遍文法は、行動の手順についての表象と解釈できるように思う。
全ての条件が出会うところ
ここまで言語誕生に必要な条件として、1)ジェスチャーやまだ言語ではない発声を通して意図を共有しようとする新しいコミュニケーション能力の誕生(背景には、自己と社会の新しい表象能力の誕生)、2)複雑な作業記憶過程に単純な音節を連合させて記憶を固定する能力(背景にはexplicit memory形成能力)、3)行動の手順を表象し表現する能力(背景には統語能力)の3条件を見てきた。もちろん他にも条件を考えられると思うが、そろそろこれらの条件が集まって言語誕生へと向かう過程について考えてみよう。
それぞれの能力がいつ発生したのかはわからないが、言語の発生時期と考えられている10−5万年前に急に現れた能力ではないと思う。おそらく、2足歩行のサルと言えるオーストラロピテクスから、人類として地上生活を始めたH.Ergasserが現れ、類人猿とは全く異なる社会生活が始まった頃から、人類はそれぞれの性質を備え始めていたと考えられる。実際この後、オスとメスの大きさの差がなくなり、噛みつくときに使う犬歯が退化する。すなわち、ボスが支配する集団から、争いの少ない平等社会に移行し、嚙みつく代わりに道具を使って骨を砕き、獲物を獲る狩猟生活に移ったと考えられる。しかし、もし多くの学者が信じているように、私たちホモサピエンスと数万年前まで交流があったネアンデルタール人や、デニソーワ人が言語を持たなかったとすると、条件は揃っていても、言語誕生のないまま200万年近い狩猟生活が続いたことになる。そして、10万年前になぜホモサピエンスがこれらの条件を現在の形の言語へと集約させることができたのか、またそのきっかけは何なのか、言語学の最も重要な問題になった。
異論もあると思うが(ネアンデルタール人も言語を持っていたと考える研究者は少ないが存在する)、条件が集まるきっかけを得ることができなかったネアンデルタール人は言語を手に入れることなく滅びてしまった。一方、我々ホモサピエンスのみが言語を手に入れることができ、地球の隅々にまで広がり支配者として君臨することになった。
ニュルンベルグのマイスタージンガー
実際きっかけについては、様々な説がすでに提案されているので成書を当たって欲しいと思う。この稿で紹介したい私の勝手な妄想は、道具(精巧な石器)作りを教える過程でこれらの条件が揃ったというものだ。そしてこのきっかけに、音楽の脳、道具の脳、そしてコミュニケーションの脳が出会いを媒介したのではないかという考えだ。これからそれについて説明するが、この可能性を考えている時、私の頭にフッと浮かんだのがニュルンベルグのマイスタージンガー第二幕のシーンだ。
オペラファンに説明する必要はないが、馴染みのない方のためにニュールンベルグのマイスタージンガー第2幕を少し説明しておこう。このオペラは、ニュルンベルグの古いマイスター社会に飛び込んできた新しい思想を持った騎士Walter von Stolzing(ワーグナー自身のことでもある)が、ニュルンベルグのマイスターからその芸術の重要性を認められるまでの「てんやわんや」を描いたオペラだ(こんな紹介は顰蹙ものかもしれないがお許しいただきたい)。孤立無援の騎士の持つ新しい芸術の可能性を最初から認め、支援するのが靴作りのマイスターHans Sachsだ。2幕ではSachsを中心に様々な場面が展開する。特に私の印象に残るのが、Walterを慕うEvaの悩みを聞いた後、Walterを貶めようと躍起になるベックメッサーが登場する前に靴を打ちながら歌うアリアだ(図2)。

図2:靴屋のマイスターハンスザックス。ニュルンベルグのマイスタージンガーの主役は騎士ウォルターではなく、間違いなくザックスだ。
ではどうして、この場面が言語誕生を想起させるのか?
もともと、オペラ、ニュルンベルグのマイスタージンガーは、様々な職人が歌を教養として身につけているという前提で成立している話だ。すなわち道具と音楽が一体化していることになる。道具と音楽が一体化していると考えても何も不思議ではない。道具を作ったり、使ったりする時はリズミックな音が出ることが多い。また、そのようなリズミックな音が、仲間同士の感情の共有に役に立っているのは、現在でも原始的な村落では見られることだ。
そして、なんといっても狩猟生活を送る人類の先祖にとって、道具作りは最も重要で、explicit memoryが要求される作業だったと思われる。さらに、それを他人に教えるとなると、explicit memoryだけでなく意図と目的の共有、利他性などそれまで人類が発展させてきた人類特有の能力が必要になる。このように道具作りは、言語誕生に必要な条件が全て揃う一つの状況になることは間違いない。
一方、音楽は現在も感情を伝え、共有するためには最も有効な手段だ。言葉を話す前の赤ちゃんは、感情のこもった発生とジェスチャーを使ってなんとか意思を伝えようとする。前出のスティーヴン・ミズン(Steven Mithen)は『Singing Neanderthal』の中で、言語が生まれるまで人類は音楽に近い発声をコミュニケーションに使っていたと考えている。この音楽の力は、例えばダビンチのモナリザを思い起こすときと、ベートーベンの運命の冒頭を思い起こす時を比べてみるとわかる。明らかに、音楽の方が思い出しやすい。これは音楽が感情と直に連合しており、
視覚表象より単純で、
脳回路の構造とフィットしているからだろう。
こう考えると、道具作りを教える時、ジェスチャーと共に、メッセージを伝えやすい音楽的発声が指示に使われたと思える。さらに、道具を作ること自体が音を発する。すなわち、explicit memoryを形成する時、連合させる単純な音が満ち満ちているのが道具作りの過程だ。当然、同じ音を、教える側、教えられる側が特定の事物の表象と連合させることができる確率は他の場合より高いと考えられる。その上、音楽を通じてより強く意図を共有する体制もできている。
言語誕生には、それまでexplicit memoryの形成のために各個体が個別に行っていた音節との連合を、偶然にしかし高い確率で他の個体と共有できる状況が必要になる。これが道具作りを教える状況に備わっていることは、以上の議論からわかっていただいただろうか。
こうして生まれた語彙が広まるのが簡単なことは、例えば有名なニカラグアの聾唖の子供達に自然発生した手話の例を見ればわかる。聾唖であることから言語世界から完全に切り離されて育った子供たちが集められて、200-400人規模で集団生活を始めることで、急速に彼らの間で通用する独自の手話が発達した。これは、一旦語彙が共有されると、それを教え合って、集団全体で共有することが可能なこと、語彙を共有する方法が確立すると、個々のコミュニケーションの現場で取り決められる語彙や文法は、瞬く間に集団で共有されることがニカラグアの手話の例から明らかになっている。このように言語は、集団で共有されることで、個人の脳内過程から解放され、集団に支えられ独自に進化することができる。要するに、1−2個の語彙の意味を共有することができれば、あとは自然に現在の形へと発展したのだろう。
なんとか言語誕生の大枠について書くことが
できたので、次回からはこの大枠に基づいて
各部分の詳細を詰めていこうと思っている。
[ 西川 伸一 ]
2回に分けて言語誕生についての私の考えを読んでいただいたので、私が言語誕生についてどう考えているのか、マイスタージンガーモデルから理解していただけたのではないだろうか。しかし細部の詳しい説明は抜けているし、結局「なんとでも言える」Just So Storyと思われてしまいそうだ。そこで今回から、このシナリオの説得力を高める為、幾つかの細部について補足したいと思っている。
このモデルでは、話し言葉中心の言語が発生する前から言語の基本能力は進化しており、サルとは異なる人類独特のコミュニケーションが可能になっていたと考えている。この点についてまず補足する。
いま現在私たちが使っている言語は、音をベースにした話し言葉と、文字をベースにした書き言葉からなっているが、文字が出来る前は話し言葉が中心だった。マイスタージンガーモデルでは話し言葉を中心とした言語が5万年前後に急速に発達したと考えており、この点についてはさらに補足する予定だが、もちろんそれ以前にもコミュニケーションのための方法は存在していた。例えばジェスチャーもそうだし、以前紹介したスティーヴン・ミズン(Steven Mithen)の言う音楽的抑揚のあるHolistic言語(http://www.brh.co.jp/communication/shinka/2017/post_000014.html)もその一つと言える。
言語の本質を考える時、話し言葉が成立する条件だけに限定すると、言語誕生に最も重要な条件を見失うことになる。この重要な条件とは、人類のみが獲得した能力で、具体的には、経験を明瞭に記憶し、記憶から呼び出した表象を、実際の対象が存在しない時に相手に伝える能力だ。
図1 シカ狩りを例に言語の条件を考える(説明は文中)
例として、複数の個体が協力して狩りをする状況を考えてみよう(図1)。何人かで狩りに出かけ獲物を探している時、目の前に鹿が現れたとしよう(設定1)。眼前の鹿の存在を皆で共有することはたやすいし、鹿を獲物にしようと全員が目的を共有することは人類でなくともたやすいだろう。人間なら、ポインティングやジェスチャーなどで標的を共有する動作が出ると思うが、ライオンのように複数で狩りをしてもほとんど個体間のコミュニケーションのない動物でも、目の前の獲物に共同して本能的に飛びかかることはできる。またその時、それぞれの頭の中に、鹿のイメージ、匂い、声などが表象され記憶されるだろう。脳科学的に言えば、このようにして生まれる鹿の表象はどの個体でも同じような脳の領域を用いて記憶されていると思う。
何回かの狩りの経験を重ねた個体では、脳に鹿の表象の記憶がだいたい同じように成立していると考えて話を進める(図中で記憶と示している)。もちろん、以前に紹介したように、人間と他の動物では前頭前皮質連合野の発達の違いでexplicit memory(鮮明な記憶:顕在記憶と邦訳されている)の程度が質的に異なっていると考えられる。しかし、ライオンでもサルでも、匂いがすれば鹿、あるいは獲物の表象を何らかの形で呼びおこしていることは間違いない。すなわち、鹿=獲物についての視覚的表象と、音や匂いの表象が連合して記憶され、新たに音や匂いの表象を体験した時、目の前に鹿がいなくても鹿の表象に結びつけることが出来る。このように鮮明度はともかく、各個体が匂いや音といった刺激により、獲物を表象する神経過程は動物から人間まで共通に備わっている。
次にこのような記憶が成立した個体が集まって獲物を探している状況を考えよう(設定2)。この時、一人、あるいは一頭の個体が匂いを通して真っ先に獲物の気配に気づくことはあり得る。獲物はまだ見えないのに、気配に気づいたとすると、そのことを伝える必要がある。でないと獲物は逃げてしまう。この時人類なら、頭の上に指を突き出して、鹿の形態を真似て鹿が潜んでいることを伝えることができる。ほとんどコミュニケーションの手段を持たないライオンは別にしても、例えば人類に近いチンパンジーの狩りのビデオを見ると(https://www.reddit.com/r/videos/comments/6wqocb/chimpanzees_hunting_monkeys_is_both_amazing_and/)、樹上のオナガザルに気づいた個体が立ち止まって木を見上げると、他のサルも止まって見上げていることから、獲物がいることを態度で示していることがわかる。このビデオではその後、一匹のオスが獲物に近づいて木の上のサルを追いかけ、逃げ惑うサルを待ち伏せしていた他のサルが仕留める様子を映している。すなわち、気配に気づいたことが習性に従って自然に態度に表れ、その態度を感知した他の個体が、学習したパターンに従って協力して狩りを行っている。このビデオでは、ジェスチャーやポインティングは全く見られない。
もちろん人間でも場合によっては、同じように決まったパターンで行動することもあるだろう。しかし、必要ならジェスチャーやポインティングを使って自分が獲物に気づいたことを伝えることができる。一方、サルは両方の手を使うことができるものの、ジェスチャーで獲物の形態模写を行うことはおそらく未だかって観察されたことはないと思う(確かめたわけではない)。
この過程をもう一度整理してみると、
1)鹿の視覚的表象と、匂いや音の表象を連合し、記憶する(人類・動物共通)
2)匂いや音の表象から、鹿の視覚的表象を呼び起こす(人類・動物共通:表象の鮮明度は人類がすぐれている?)。
3)鹿の視覚的表象を、ジェスチャーで形態模写し、それを他の個体も理解する(人類特有)。
という一連の過程が起こっている。とすると、最後の形態模写ができる点が人類のみに備わった言語誕生の条件になる。すなわち、臭いという指標で活性化された視覚イメージが視覚野を興奮させるところまでは他の動物も同じだが、その表象を形態模写しようという着想と、実際に表象に基づいて形態模写する行動は人間特有になる。もちろん、表現にジェスチャーを用いる必要はない。コミュニケーションという目的なら、鹿の声を真似てもいいし、今匂いを感じていることを、鼻を広げて表現してもいい。要するに、頭の中に呼び出した表象が、他の個体の脳にも誘導することがポイントになる。チンパンジーを見ていると、狩りの時両手が使えると言っても、実際にジェスチャーで何かを表現することは解剖学的に難しいと思う。また、鹿の声を真似ること同じ理由で難しい。このように、新しいコミュニケーション手段が発達した背景には、もちろんこのような解剖学的な差も存在する。
この時、ツノを突き出す形態模写は実際の鹿のイメージに近く、セッティングにもよるが表象(意味)を他の個体と共有できる可能性は高い。パース流に言えば(http://www.brh.co.jp/communication/shinka/2016/post_000012.html)、イコンを用いる表現によるコミュニケーションになる。
一方、鹿の声を真似て表現するのは、鹿そのものを表現しているわけではないが、声が鹿の存在を示し、また鹿の表象を他の個体の脳に呼び起こす指標になると言える。これは、パースの言うインデックスと言えるが、話し言葉のルーツと言えるのかもしれない。
鼻をクンクンさせて、今匂いがしていることを伝え、他の個体も同じ匂いを嗅いでみて確かに鹿だと確認することもできる。この場合は鹿の匂いというインデックスを共有したいことを、「嗅いでみろ」と形態模写で伝えることで、最終的に同じインデックスを相手にも嗅がせて、鹿の表象を他の個体の脳内に形成させることになる。このように、コミュニケーションの手段は言葉がなくとも多様に存在するが、いずれの方法でも、最終的に自分の頭の中に呼び起こした鹿の表象を、他の人の頭の中に呼び起すことがゴールになる。
ではこの過程を支える人間特有の能力とはなんだろう。
すでにゴールを他の人間と共有して協力関係を樹立できるのは人間特有の能力であることを述べたが、獲物の表象を共有する過程を考えてみると、言語の背景にも同じ能力、すなわち
1)ゴールを共有する協力を可能にする人間特有の能力、
2)explicit memoryの形成能力、
が存在することがわかる。この能力の脳科学的背景として、ホモ・エルガステル(Homo.Ergasser)の誕生から始まる、前頭前皮質の連合野の急速な発達があることは間違いない。重要なことは、これらは全て学習により発生するのではなく、自然に発生する能力である点だ。explicit memoryが脳の様々な領域を連合させる前頭前皮質の発達に強く依存していることはわかるが、ゴールを他の個体と共有して協力する能力が、連合野がどの領域と結合することによるのかを特定することは現段階では難しい。
さらに言語は進化し、最終的に話し言葉(音節)をシンボルとする段階に至るが、全て上に述べた最初の言語へ向けた脳内のネットワークに起こった質的ジャンプの延長として考えることができる。鹿を求める狩りを例に、さらに進んだ内容のコミュニケーションを考えてみよう。
例えば昨日鹿を見た場所に今日も行ってみようと相談をするという設定が考えられる(設定3)。この時、実際の鹿が近くにいて気配を感じているわけではない。従って、具体的な鹿の存在にまつわるイコンやインデックスは存在しない。すなわち、この時鹿について語るためにジェスチャーや鳴き声を真似ることは、狩りをするという目的が共有されている必要がある。そしてこの場合、この目的が記憶から鹿の表象を呼び起こす刺激として働き、目的を共有する他の個体にも同じ刺激として働いている。すなわち、存在しないものを表現できている点で、我々の言語に極めて近いレベルに達している。そしてこのためには、レベルの高いexplicit memoryと目的の共有が必要で、これは人間だけが達成した能力だ。残念ながら、何か新しい分子の進化に基づく質的変化かどうかは明らかではない。
このように実際には存在しない表象を共有するための記憶が成立し、目的のような抽象的な表象が共有できる段階に達すると、ジェスチャーや鳴き声といったインデックスにより呼び起こされる表象はすでに脳内の様々な表象と連合している。例えば、狩りのプランに実際に必要なのは、肉にありつくため獲物を得ることで、目的はジェスチャーで表現した鹿の表象に限らない。以前紹介したように(http://www.brh.co.jp/communication/shinka/2017/post_000022.html)、私たちの脳内の表象は決して単独で存在するのではなく、様々な表象とカテゴリー化して連合している。鹿の表象は獲物になるほとんどの動物と連合しているだろうし、また獲物以外の危険な動物など多くの記憶とも連合している。実際、狩りで最初にイノシシに出会っても、プラン通りアタックすることになるだろう。
以上をまとめると、言語能力の条件として、
1)情報を共有したいという強い欲求、
2)レベルの高いexplicit memory(高い表象間の連合能力)
3)実物に依存しない表象の呼び起こし
4)これらをストーリーに仕上げる統語能力に必要な、身体的行動の表象能力
があり、これが揃えば話し言葉も目の前だと考えている。
確かに、最後のセッティングでは目的と言った抽象的なきっかけで異なる個体が同じ表象を共有するなど、かなり高度に見えるが、条件自体は話し言葉を持たない直立原人や、ネアンデルタール人も持っていても何の不思議はないと思っている。しかし、コミュニケーションの為のボキャブラリーを増やすには、音節をシンボルに使う、話し言葉の誕生が必須だった。
これが起こったのを、マイスタージンガーモデルでは5万年前と推定しているので、次回は、我々ホモ・サピエンスの歴史を見ながら、5万年前に現在のような話し言葉が始まったと考える根拠について補足する。
[ 西川 伸一 ]
前回述べたように、言語発生に必要な脳の条件は現生人類誕生よりずっと以前に整っていたと考えている。ただ、現在私たちが日常使っているような言語、すなわち話し言葉を基盤とする言語(ここではspeech language:S言語と表す)となると話は別だ。すでに議論したように、S言語では脳内に表象される対象と何の関連もない音節がボキャブラリーとして対応し、しかもその対応が異なる個体間で共有される。これまで述べてきたように、これが可能になることで、無限の表現力が我々に備わった。しかし、おそらく小難しい議論を必要としない狩猟採集民の生活には、ジェスチャーや、一種の赤ちゃん言葉の発声で十分間に合ってたはずなのに、S言語への指向性が生まれるにためは、より高いレベルのコミュニケーションが要求される状況があったと考えられる。
「マイスタージンガーモデル」では、複雑な道具作りを教えるという状況で生じた様々な必要性がS言語誕生を促したと考えている。今回の補足では、これが5万年前に起こったとなぜ考えているのかについてその理由を説明してみたい(断っておくが、これはあくまでも個人的な意見だとしてお読みいただきたい)。
現生人類の発生と移動
先に答えから明かそう。私が考える5万年という数字の由来は、我々の先祖ホモサピエンス(サピエンス)がアフリカで誕生した後、ヨーロッパへの進出を始めた時期から算定した数字だ。すなわち、この時期にS言語が初めてサピエンス、特にシナイ半島に住んでいたサピエンスに誕生したのではと想像している。なぜS言語の誕生とヨーローッパへの進出が重なるのか?
これまでに発見されたサピエンスと思われる最も古い骨は、ライプチヒ・マックスプランク人類進化研究所の研究者たちによりモロッコIrhoud から発見され、31万年前の骨と特定された(Hublin et al. Nature 546:289, 2017)。この発見により約70万年前にネアンデルタール人(以後ネアンデルタール)からアフリカで分離したサピエンスが、南、東アフリカだけでなく、モロッコの位置する地中海の西の端までアフリカ中に広く分布していたことが初めて確認された。すなわち、人口が増え、繁栄を遂げていたと思われる。
2つの出アフリカルート
こうしてアフリカの隅々に分布したサピエンスは、15万年前にアフリカを出て、まずアジア、オセアニア方面に移動を開始する(図1)。
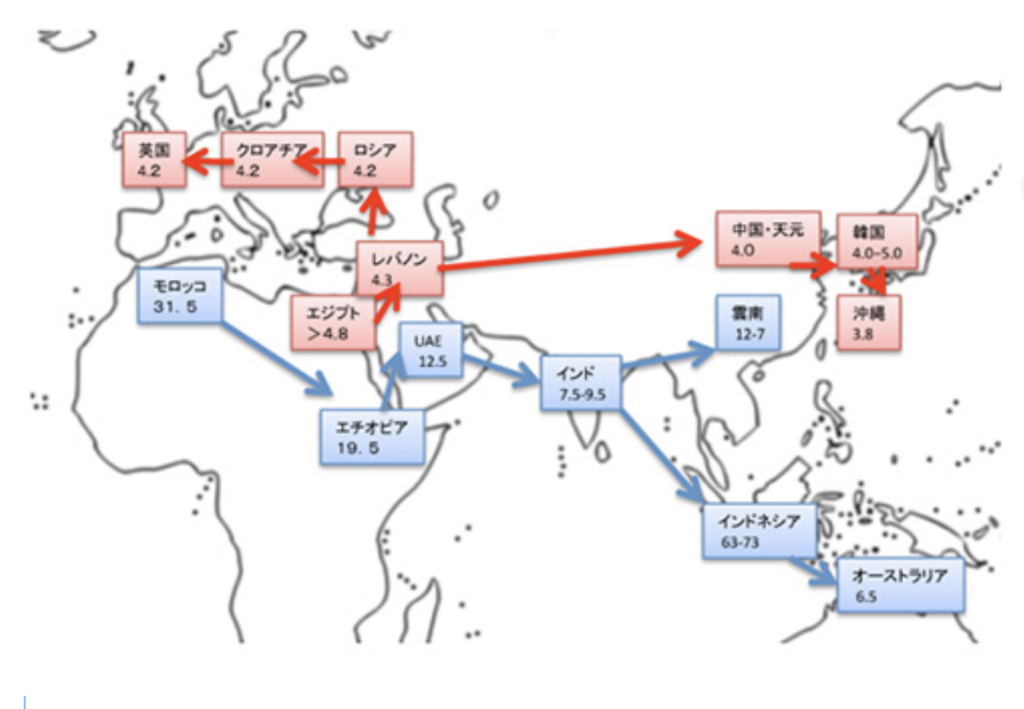
図1: 『Science』に掲載された総説をもとに(Science 358: DOI: 10.1126/science.aai9067, 2017)筆者が作り直したもの。サピエンスと確認できる遺跡が存在する場所とその年代をつないで移動ルートを割り出している。
図1はサピエンスがユーラシア進出にたどった2つのルートを示しているが、ホモサピエンスによる可能性が高い遺跡を古い順につないでいくと、このルートが見えてくる。中でも最初のステップ、すなわち出アフリカだけに焦点を当てると、両方のルートでそれほど時間差はない。アフリカ以外で見つかる最も古いサピエンスの骨はイスラエルで約16万年前のもので、アラビアルートとほぼ同時期か、あるいはより古い時期のサピエンスだ。すなわち、両方のルートでサピエンスの出アフリカが15万年前には始まっていたと考えられる。
最近『Nature』に、30万年前ぐらいの地層から、直立原人由来とは考えられない進んだ石器が出土したことが報告された(Nature 554:97, 2018)。この石器がサピエンス由来で、ネアンデルタール人やデニソーワ人でないとする証拠はないが(人骨が発掘されていない)、明らかに著者らは、サピエンスが定住はしていなくても、インドにかなり早くからに到達していたと結論したそうに思えた。今後発掘が進むと、アラビア・インドルートは早くから完全に開いていたとする結果が出てくる可能性があるが、現在まで得られている最も確実な証拠に基づくとサピエンスのアジアへの移動は15万年前後に起こったとするのが適切だろう。
さて、15万年前は地球がかなり冷えていた時期でおそらく現在より6度以上温度は低かった。しかしその後急速に温暖化が進み、13万年前にはほぼ現在と同じような気候になったと考えられる。この急速な温暖化は、13万年前から急速にサピエンスがアジアに移住を始める一つの理由になったと思われる。その後、オーストラリアには6.5万年前に到達しており、インドを経由するサピエンスの東進を阻む他の人類はいなかったと思われる。
閉ざされたシナイ半島ルート
ところが、図1を見ていただくと地球が温暖化して出アフリカが加速した時期にも、シナイ半島からヨーロッパへのルートは5万年前まで全く閉ざされていたことがわかる。図2は、図1に示した地図状にネアンデルタール人の分布を筆者が書き加えたものだが、サピエンス進出のヨーロッパルートとネアンデルタール人の分布が一致する。異論もあるとは思うが、シナイ半島ルートが5万年まで、全く開かなかった理由は、ヨーロッパルートに分布していたネアンデルタール人が、サピエンスのシナイ半島からの北進を阻んでいたのではないかと想像できる。
図2:図1にかぶせてネアンデルタール人の分布を重ねてある。
先にも述べたが、アフリカ外で発掘された最も古いサピエンスの骨は16万年前のイスラエルカメル山近くで発見された骨だ(Hershkovitz et al, Science 359: 2018)。すなわち、シナイ半島ルートは、アラビア・アジアルートと同じくかなり早い時期にサピエンスが進出したにもかかわらず、ネアンデルタール人の存在により10万年以上もの間、行く手を阻まれていたと考えられる。
図3『Nature』 に掲載された記事(Mellars et al.Nature 479:483, 2011)をもとに、筆者が作り直した図で、サピエンスのヨーロッパ進出の3本のルートを示している。
ではなぜ5万年前にこの均衡が破れたのか?
私自身はこの理由を、話し言葉を基盤とする言語(S言語)がシナイ半島でサピエンスだけに誕生したからと考えている。
サピエンスのヨーロッパ征服の理由
まず図3に示した、ヨーロッパへのサピエンス進出を見てみよう。これまでヨーロッパ内で発見され、ゲノム解析がほぼ完全に行われたのは、ルーマニアで発見された約4万年前のサピエンスの骨で、なんと6−9%のゲノムがネアンデルタール人由来であることがわかっている(Fu et al. Nature 524:216, 2015)。このことは、サピエンスのヨーロッパ進出が、ネアンデルタール人と常に接しながら進んだことを物語っている。さらに、4.4-4.2万年前にサピエンスがイギリスやスペインにすでに達していたということは、かなり短い期間でヨーロッパ全土の征服を成し遂げたことになる。
この背景として、1)ネアンデルタール人の生存が特異的に脅かされる自然要因、2)ネアンデルタール人との交雑によるサピエンスのヨーロッパへの適応、そして3)サピエンスに起こった文化・技術の大きなイノベーション、などが考えられる。
自然条件についてみれば、5万年以降地球は温暖と寒冷の間をめまぐるしく行き来した時期だ。また、7万年前には現インドネシア・トバ山の大噴火、また4−5万年前にはイタリア・ナポリにあった火山の大噴火があり、ヨーロッパの気候はめまぐるしく変化したと考えられる。これによる食物の減少は、ネアンデルタール人の住む北部でより大きな影響を与えたと考えられ、サピエンスの進出を促した可能性は否定できない。
ネアンデルタール人との交雑により、サピエンスが寒冷地で生存できるよう適応した可能性もある。事実、我々現代人も、ネアンデルタール人から寒冷地で生きるための様々な遺伝子を受け継いでいることが明らかになっている。ただ、サピエンスとネアンデルタール人の接点で暮らす人類にネアンデルタール人遺伝子が高い比率で流入しているとすると、なぜ10万年以上両者の均衡が崩れなかったのか理解しがたい。おそらく、この要因はヨーロッパ征服にそれほど貢献していないのかもしれない。
こう考えてくると、結局5万年前のサピエンス優位は、文化的、技術的な要因が大きいと考えられる。要するにサピエンスがネアンデルタールには真似できないイノベーションを成し遂げた結果、それまで続いた均衡が大きく崩れたという可能性だ。またこの差が、そのままネアンデルタール人の絶滅に続いていくことになる。
ネアンデルタール人とサピエンスの差
事実、ネアンデルタール人がサピエンスより劣っていたと考えている研究者は多く、
Wagnerのオペラ「ニーベルングの指環」では、槍や刀がぶつかりあって壊れるという場面が何度も出てくる。例えば鍛えられた強い剣でも人間(ジーグムント)が構えたノートゥングは、神(ヴオータン)の槍にうち砕かれる。これは武器の優劣を示すことで、神の力を示そうとしていると思うが、ともすると私たちはネアンデルタール人と現生人類の間に同じような関係を想像してしまう。しかし前回も述べたが、武器や戦略の差が両者の絶対的な優劣の差の先行原因だとは思いにくい。
事実、ネアンデルタールと現生人類はシナイ半島では10万年近く隣接して暮らしながらも均衡を保っていた。シナイ半島および北上したばかりの現生人類は、使っていた石器(ProtoaurignacianやAhmarian文化とよばれている) でネアンデルタール人(Levalloi文化)から区別されている。しかし、最近の研究でシナイ半島や現生人類が北上したルートではこの区別が明確でない事もわかっている(Hublin, PNAS 109, 13471, 2012)。則ち、石器など道具が優劣の決定的要因になったとは考えにくい。
そこで前回この優劣の鍵となったのが、VerbalあるいはSpeechを媒体とする言語(S言語)であると提案した。S言語獲得により優劣がハッキリした後は、現生人類でさらに大きな文化的変化が起こり、おそらく石器自体もAhmarianやProtoaurignacianからその後急速に発展するきっかけとなったと思われる。
では、なぜわざわざ石器作りを教える過程がS言語誕生の現場となったと考えるのか?今回はこの問題を考えてみるが、せっかくの機会なのでまず石器作りについて勉強する事にしよう。
石器作りに関する本はおそらく他にも数多く出版されているとは思うが、私の知識はもっぱらジョン・シー(John J Shea)著の『Stone tools in human evolution:behavioral differentces among technological primates』 (Cambridge University Press)に頼っている(図1)。ほかの本を読んでいないので、これだけを推薦というわけには行かないが、考古学の論文を読むときにはきわめて役に立つ本だ。勿論ここで述べる事も殆どがこの本を参考にしていることを断っておく。

図1 John J Sheaの著書。
石器について人間だけでなく、サルが使用した跡についての区別の方法にいたるまで書かれている。通読すると、石器が如何に奥の深い話かわかる。
出典:Amazon co.jp
人類が石器を使い始めたのは300万年以上前のことだが、石器は人類の進歩と共に発展してきた(図2)。この歴史はまず旧石器時代と中石器時代に分けることが出来る。中石器時代は、だいたい1万年ぐらいから始まっており、既に弓矢など高度な武器が発達している。私達が問題にするのは、それ以前の旧石器時代で前期、中期、後期に分けられている(図2)。

図2 石器の発達。Sheaの著書を参考にして、筆者が作り直した。
写真の出典:Wikipedia
各時期の石器は代表的発掘場所の名前がついており、そこで発掘される骨の形状から、使用していた人類がわかっている。ただ、示している以外の人類が使っていた可能性を排除することは出来ない。
この発達の歴史はまさに石器の作り方の開発の歴史だが、殆ど二足歩行のサルと言っていいアウストラロピテクスと、常時2足歩行の直立原人の間で大きな変化が起こっているのが図2からわかる。すなわち、直立原人の登場が、様々な点で人類最初の大革命をもたらしたと考えられる。
実際、実験的に石器を作成する実験考古学によると、Oldowan文化では、拾った石を割って、そのかけらをもう一度叩いてみることぐらいしかおこなわれていない。その後、直立原人によるAcheulian文化がはじまると、急速に石器製作の工程が質量ともに増加し、複雑度が増していく。この結果、後期Acheulian文化では、1)石器に適した石の採石、2)砕石、3)破片の選択、4)破片の加工、5)加工する対象の最終的選択、6)様々な調整による形作り、などかなり複雑な過程が石器作りに使われていたことがわかる。おそらく、実際に使用した後の使い勝手についてのフィードバックも行われただろう。すなわち、デザインの構想とそのための加工技術の発達が石器の発達を支えたと言える。
このように、石器を見ていると、直立原人が既に高度な文化をもっており、石器製作に必要なExplicit memoryやゴールを共有して協力する能力、すなわち言語能力を十分備えていたことを実感する。
石器作りでもう一つ重要な点は、このような工程を、個人が個別にやっていたというより、一種の工房でまとまって行われていた点だ。このような工房で石器や他の道具を作り、後進に教えるという作業は、当然高いコミュニケーション能力が前提となる。すなわちこの能力が高さに応じて、石器の機能が発展したと想像される。
これらの点を考慮すればするほど、この工房で石器作りを教え、また習おうとする過程こそが、音節を単語として用いるS言語の発生場所に違いないと私には思えてくる。その理由をさらに説明しよう。
理由1:音はexplicit memoryに必要な連合を助ける。以前「コミュニケーションと言語」で述べたように、おそらく人類は、重要だが複雑なイメージを記憶する際、単純な表象と連合させることを自然に行っていたはずだ。この時、常にまわりで繰り返して聞こえてくる音節は、連合相手として利用価値が高い。
理由2:石器作りの現場では様々な繰り返す音で満ちている。これについては想像してみるだけで十分だろう。先にまとめた過程で石器を作ろうとすると、各過程で違った音が発生し、それも繰り返して何度も聞こえる。
理由3:石器作りの現場には表現するべき具体的対象が多く存在する。石器作りには多くの工程が必要だが、そのおかげで現場にはその工程を代表する具体的なモノ(・・)が存在しており、複数の個人で同一のモノ(・・)を対象として共有しやすい。
理由4:石器作りを教え、教えられる関係には、個人的な表象を他の個体に共有させる階層性がある。個人が、自分の頭の中で特定の音節を複雑なイメージの表象として連合させることは特に難しい話ではない。しかし、個人的な表象を、他の個体と共有するためには、表象されている具体的モノ(・・)を先ず共有することが必要で、それを共有した上で、誰かがそのモノに対応する(表象する)音節を提示し、他の個体にもモノ(・・)と音節の関係を共有してもらう必要がある。理由3に述べたように石器作りの現場にはこの具体的モノ(・・)が存在し、指差しでそれを共有できる。
その上に、教える側と、教えられる側の階層が存在することも重要だ。具体的モノ(・・)を前にしても、それぞれが勝手な音節を提案するのでは収集がつかない。しかし、教える側がはっきりしている場合は、主導権を教える側が持って、モノ(・・)と音節を関連づけ、相手に共有させることが可能になる(図3)。

図3 具体的なモノの共有、上下関係、様々な音がある現場でこそモノと音節の対応が共有される
イラストの出典:無料サイトPixabay
理由5:石器造りで発生する音は時に音楽的だ。道具を作る過程で出る音を音楽に取り入れる能力でワーグナーは天才と言えるが、道具作りも手馴れてくると、自然にリズムに合わせて工程を進めることも行われただろう。音楽が感情を表現し伝える手段だとすると、道具を作る時の音を聞きながら、音楽を思わず口ずさむこともあっただろう。
一方、音節が中心の言語が生まれるまでは、コミュニケーションにはジェスチャーとともに、スティーヴン・ミズン(Steven Mithen)のいうような音楽的フレーズも感情を伝えるために使われたと考えられている。例えば「オーケー」とか「グーーーッド」とうなづくような感じだ。とすると、この現場には声もあふれていたと思える。
もともと、石器作りを教えるためには、高いレベルのコミュニケーションが必要になる。このため、S言語発生前から、音楽的発生でのコミュニケーションを図ろうとする努力は日常的に行われていたと思える。
理由6:石器作りの現場には自然に文法が生まれる。石器作りでは、プランに基づいてどう身体を動かすかが問題になる。既に文法のルーツの一つは身体の動きの手順だと提案したが、その意味で石器作りを教える過程で音節を使った単語が生まれたとすると、それを並べる統語も自然に生まれたのではと考えられる。
以上、私が石器作りを教えていた工房こそが、S言語発声の現場だと考える理由について述べたが、全て私の妄想として聞いておいていただきたい。
大事なことは、このように音節を用いてコミュニケーションが図れることが理解されると、S言語は急速に仲間に広がることだ。これは、すでにニカラグアのろうあの子供達の手話が共有されていく過程の解析から明らかになっている。最初、2−3人の少人数の中で生まれたサインが瞬く間に集団全体に広がり、今度はあらゆる場所で新しいサインが作られ、広がっていく。
手話と比べた時、音節を用いるS言語は、表現力だけでなく、表現のスピードでも高い能力を発揮する。この便利さが一旦認識されると、石器作りの現場以外でも新しいボキャブラリーを形成する過程が進み、集団で共有するボキャブラリーは急速に拡大できると考えられる。
この結果、より複雑なコミュニケーションが可能になれば、もちろん優れた石器を開発する能力も高まるだろう。しかし石器の機能に差がないとしても、音節を単語として複雑なコミュニケーションを迅速に行える能力は強い武器として、それを持った集団と、持たない集団の間には、大きな優劣が生まれるはずだ。
もちろん、それまでのジェスチャーや単純な発声だけでは表現できない高いコミュニケーション能力を手にしたいという必要性と、ここに述べたような条件が揃えば、S言語の誕生は石器作りを教える現場である必要はない。
最後にもう一つ妄想を述べると、同じ現場では、絵を使うコミュニケーション手法も誕生しただろう。実際、多くの職人さんも、技術を伝える時、絵を使うことが多いと思う。とすると、石器作りを教える現場は、S言語とともに将来の文字誕生のきっかけになったのかもしれない。
これでようやく言語誕生について終えることができる。次から、言語と文字の統合の問題をかんがえる。
[ 西川 伸一 ]
言葉の流れに対する脳の反応
2018年4月2日
言語の誕生については前回で終わり、今回から文字(Writing)について考えようと準備していたが、たまたま最近、連続的に聞こえてくる言葉の流れに対する私たちの脳の反応についての面白い研究を読んだので、予定を急遽変更し、今回はこの問題を取り上げたい。
言うまでもなく言語は私たちの脳活動から生まれた情報メディアで、個人の脳のネットワークに集め、記憶し、処理した内部・外部からの情報を、ほぼ同時的に他の個体と共有できるようにして伝達することを可能にした点で、それ以前に生物活動から生まれたDNAを含む情報メディアとは全く異なっている(言語の起源I参照)。しかしこの情報メディアは、完全に脳の活動に依存しており、言語情報は他の人間の脳回路へと集められ処理されてはじめて情報として機能する。すなわち、私たちの脳が音として流れてくる言語を聞いた時、音節で区切られた単語が集まった文章として表象された情報に注意を向け、その表象を脳内に長期記憶として持っている様々な表象と参照しながら、脳回路上の新しい表象へと転換しなおすことで、情報を理解している。従って、言語を聞いて、情報として脳内に新しく表象し直すまでの過程を知ることは、人間の脳科学に残された重要な領域だ。
言語誕生以来、私たちは言語をそれが実際に使われているのを聞いて経験する中で獲得してきた。これは学校で何かを論理だって習うというのとは違っており、単語とその時の経験を連合させる過程を繰り返しながら、単語の意味を理解し、ボキャブラリーを増やす、一種の記憶過程だ。早い遅いはあってもほとんどの子供が3歳前後でかなりの数のボキャブラリーを習得し、言語を自発的に話せるようになるのは、言語自体が人間の脳の一般的発達過程にうまく適合していることを示しているのではないだろうか。
これは、言語か各個人によって別個に習得される必要性があることから、言語自体もこの習得過程に適するように進化して来たと考えるとわかりやすい。この言語の特徴を、以前紹介した(http://www.brh.co.jp/communication/shinka/2016/post_000011.html)Deaconは、昔アップルコンピュータがアイコンを用いた画面を使って、PCオペレーションを私たちユーザーの脳に合わせるという革命を成し遂げたことにたとえている。この例えを使うと、PCが私たちの脳に合うように進化したのと同じように、私たちが経験を通して学びやすいように言語も進化しているというわけだ。
ただ、この考えが本当かどうかを判断するためには、脳が言葉の流れとして表象されている情報を、どのように処理し、脳回路を書き換えるのか理解する必要がある。現在の言語は我々の脳にフレンドリーだとしても、聴く前から情報の内容がわかることはない。順序だって並ぶ単語を一つづつ聞きながら、全体の情報の意味をどこかの時点で理解する必要がある。
この時成人の脳でどのような過程が起こっているのか想像すると、一つ一つ流れてくる単語を、作業記憶として処理しながら、意味を理解するために作業記憶を長期記憶や、意味記憶と常に参照しながら、情報をまとめ上げていると想像できる。長期記憶や意味記憶が脳内の様々な場所にバラバラに蓄積されているとすると、言葉を聞いて理解する過程は、多くの脳内領域が作業記憶と参照される脳全体の活動だとわかる。問題はこの過程に対応する脳過程をどう観測するかだ。
これまで脳の活動を記録する様々な方法が開発されているが、PETやMRIは脳の興奮に伴う血流の変化を調べる検査法であるため、実際の活動と、検査上の変化とどうしても1秒以上のズレがあり、流れてくるそれぞれの単語に対する反応をリアルタイムで調べることは難しい。このため、どうしても脳の電気活動を調べる方法に頼ることになる。研究者側から見て一番望ましい検査は、脳内に電極を電気活動を拾うことだが、おいそれとできる検査ではない。結果として、いわゆる脳波計(EEG)を用いる検査が行われるが、痒い所を「布団の上」から搔く様な話で、頭蓋の外から記録される電位変化の解釈は難しい。しかし、他に簡便な方法がない以上、EEGを出来るだけ多くの領域から記録し、得られる波形をコンピュータを用いて解析する方法が進んだ。特に、時間的に連続して生起するイベントに対するEEGの変化をEvent related potential(ERP)と呼び、小さな波形の変化を抽出して解析する方法が進んだ。
そして1980年、この分野で最も影響力の高い論文がKutaとHillyardによりScienceに発表された(Science 207:203, 1980)。 この論文で示されたのは、文章を聞いている時、急に文脈に合致しない単語が紛れ込んでいると、その単語を聞いてから約400ms後にEEGの波形が明らかに変化するという現象で、N400としてこの分野で最も重要な現象として研究されている。
彼らの総説から例を挙げてみよう。「彼は毎日ヒゲと町を剃る」という文を聞いた時、文章の中の町という単語は場違いであるとだれでも思う。実際、脳の方も困惑しているのか、「町」という言葉を聞いた400ms後に電位が高まるのを観察することができる。これが、N400の発見だ。
N400はその後40年近く、文章として表象される言語に対する脳の反応を検査するための重要な指標として研究されてきた。個人的な解釈を述べるのを許してもらうなら、N400とは聴きながら作業記憶として処理する単語やその並びを、これまで脳内に形成した長期記憶と参照しながら、一つの情報へとまとめ上げる作業過程と言える様に思える。そして、文章を聴き終わるまでに、私たちは文脈を理解しており、このため逆に場違いな言葉が紛れ込むと、強く反応してしまうことになる。
考えてみると、3歳までの発達期で私たちは場違いな単語と出会うという経験を繰り返してきたはずだ。単語自体は抽象的で何の実態もない。したがって、脳内の長期記憶と対応付けられないと、全ての単語はその場に合致していないことになる。とすると、発達期に耳にする単語は全て場違いな単語で、長期記憶と連合できないという点で、意味をなさないはずだ。しかし、音と具体的な情景や物が繰り返し連合することで、単語とともに言語としての認識が、長期記憶として脳に蓄積されていく。そして、言葉を聞くたびに、外界の表象とともに、言語として蓄積した長期記憶が参照される。これが、N400として記録される過程ではないだろうか。
面白いことに、N400は例えば音楽を聴いていて不協和音や雑音を聞く時には観察されない。このことは、音楽と言語の把握が大きく異なるメカニズムで行われていることを示すとともに、N400が、言語意味が問題になるEPR過程に限定されるプロセスである過程を強く示唆している。N400という現象論が、より実際の経験や理解に転換するためには、これまでとは異なる新しい方法が必要になるだろう。しかしEEG検査は幼児期から可能な検査であることを考えると、今のままでも、発達段階で言語体験とN400を調べることで、言語についてさらに新しい発見がある予感がする。
このようにN400は奥の深い研究領域で、完全に理解されているわけではなく、これ以上説明するのはやめるが、とりあえず概要を理解してもらったところで、今日紹介したいと思っているのが、私がN400のことを知るに至ったダブリン大学からの論文で、N400をヒントに、言語理解の新しい展開を模索している。タイトルはElectrophysiological Correlates of SemanticDissimilarity Reflect the Comprehensionof Natural, Narrative Speechで雑誌Neuron 5月号に発行予定だ。
N400はたしかに言語に関わる過程の研究指標としては優れている。ただ、場違いな単語を滑り込ませるなど、これまでどうしても自然な言語からはかけ離れた人為的なセッティングで研究が行われてきた。
この研究の目的は、これまでのような人為的設定ではなく、自然な話し言葉の中で個々の単語に対するN400に相当する脳の反応を調べるための、新しい方法の開発だ。脳の記録自体は、EEGを用いる点で特に代わり映えはしない。また、脳波の小さな変化の中からN400に相当するコンポーネントを小さな波形の変化の中から取り出すこ方法も今の方法で十分だ。しかし、自然な言語に対する脳の反応を調べるためには、私たちが頭の中で行っているのに近い形で、それぞれの単語の間の関係性を評価する必要がある。例えば、机とりんごは大きく違っているが、りんごとみかんは近いといった関係だ。
このために、著者らは文章の中に出てくる単語の持つ400種類もの性質を元に400次元空間上のベクトルとして定義し、この値を基礎に個々の単語の持つ違いを、この空間上の距離として数値化した。この方法の詳細を理解しているわけではないが、人間が恣意的に単語同士の距離を決めるのではなく、各単語の使われ方を基盤にして、単語間の違いを独立に決めたところが重要で、これにより初めて独立した言語と、人間の脳の反応の相関を数値として調べることが可能になる。400次元と聞くと難しそうだが、おそらく自動翻訳などでAIを使って単語間の近親性を計算しているのと同じことだと思う。私たちは意味と言うと、すぐに自分の経験で考えるが、AIが発達した今は、このように多くのパラメーターを使った多次元空間での距離で定義することは普通に行われていると想像する。
様々な文章を聞かせて、EEGを記録し、文章に含まれる単語とEEGの反応の相関を調べていくと、単語自体ではないが、続いて現れる各単語間の距離と、200-500msで現れる電位とが高い相関を示すことが明らかになった。これをtemporal response function(TRF)と呼び、指標としてEEGを解釈することで、独立に定義した言語空間と脳の反応の相関が生まれる脳のメカニズムを探っている。
この論文の最も重要な発見は、ここで定義された単語同士の距離は、ランダムに単語を聞かされても、TRFを誘導しないことだ。例えばone fishと聞くときもちろん意味を理解することができるが、oneとfishの単語としての意味論的差異は大きい。驚くことに脳波、このone vs fishの違いにしっかりと反応し、fishと聞いた後300-500msでTRFの低下がみられる。すなわち、文章内の単語間の意味の違いの大きさに脳波がしっかり反応している。
しかし、同じ単語の組み合わせでも、同じ文章を逆さまに読んで聞かせた時にはTRF決して現れない。また、意味のある文章でも、周りのノイズにより理解が邪魔されると、やはりTRFは現れない。逆に、文字を見ながら文章を聞かせて理解を高めると、TRFがさらに明確になる。
これが結果の全てだが、単語間の距離という文章を構成している部分が、文章全体の文脈の中で連続的に評価されていることが大変よくわかる。この文脈を私たちが長期記憶として持っている言語空間と参照されることで形成されていく意味と捉えるとN400やTRFがこの400msの間に起こっている過程を反映しているのは納得できる。
以前言語の2重構造について述べ、言語は個人の空間と、個人とは独立した社会が形成する言語空間をもち、それぞれは相互作用しながらも、独立して言語を進化させることを述べた(言語の2重構造参照)。私は、今回紹介した論文を読んで、この2種類の言語空間を脳科学的に扱えることを確信した。今後も是非この分野に注目して行きたい。
ちょっと脱線したが、次回からは文字について考える。
[ 西川 伸一 ]
神経ネットワーク情報システムの発展
2017年2月1日
神経系もゲノム進化の産物だし、脳を含む神経ネットワークの発生には、ゲノムの指示と一体化して展開するクロマチン情報の変化が必要だ。当然、「心は身体に依存している」と同じ意味で、神経ネットワークは身体に依存し、またゲノムやクロマチン情報に依存している。それでも前回述べたように、膜電位の脱分極の伝搬による興奮の伝達および、シナプスを介する方向性を持った細胞から細胞への興奮伝達という原理を共有する細胞のネットワークは、作動原理から見たとき、身体、ゲノム、クロマチン情報など、神経系誕生前に生物を支えてきた情報システムから大幅な自由度を勝ち得ている。ともすると私たちが簡単に心身2元論の罠に落ち、「心と体」を分けて考えるのも、この神経系の独立性のゆえんだろう。
この様に、独自の原理を持ち、他の情報から大幅に独立した神経ネットワークは、それに参加する神経細胞数さえ増やせば、ほぼ無限に解剖学的・生理学的に複雑化することが可能で、事実これまで質的にも、量的にも複雑化する方向に進化を続け、カンブリア大爆発、そして現在の地球上での人間の繁栄の原動力となってきた。そこで、この過程で身体から大幅な独立を果たした神経系に加わった幾つかの重要な性質について順に考えてみたいと思う。
この進化が到達した地点は例えば次の文章を読めばよくわかる。
「星野温泉行のバスが、千ヶ滝(せんがたき)道から右に切れると、どこともなくぷんと強い松の匂いがする。小松のみどりが強烈な日光に照らされて樹脂中の揮発成分を放散するのであろう。この匂いを嗅ぐと、少年時代に遊び歩いた郷里の北山の夏の日の記憶が、一度に爆発的に甦って来るのを感じる。」
寺田寅彦の随筆「浅間山麓より」(青空文庫掲載)の一節だが、私たちは森の緑を目にし、松の匂いを感じたとき、遠い少年時代の思い出を鮮明に呼び起こすことができる。
いうまでもなく、ここに書かれた現象は全て神経細胞同士の結合が可能にしている過程だ。
今回は、このような地点に到達するまでに神経ネットワークが獲得してきた幾つかの重要な特徴について考えてみたい。ただ、神経や脳の特性について詳細な説明をしようと思うと、まさに脳科学の教科書を書くことになり、私の任ではない。ここでは、神経系を情報の観点から整理しなおすとき、私が押さえておきたいと思う点について、独断を交えて短くまとめてみようと思っている。
物理刺激の受容
私たちは、視覚、触覚、聴覚、温覚、嗅覚、味覚の6種類の感覚を持っていおり、どの刺激であれ感覚細胞が共通に使っている陽イオンチャンネルは原核生物から存在する。また、光合成に見られる光を化学反応に変化させる仕組や、鞭毛などの運動メカニズムに見られる仕組みは、光や圧力といった物理的刺激の受容に準備が整っていたことを意味している。
以前「神経細胞の誕生」について説明した時にも議論したように、(http://www.brh.co.jp/communication/shinka/2016/post_000021.html)、視覚、触覚、聴覚、温覚など物理刺激を受容できる能力の獲得は、自己と関わる環境の範囲を大きく拡大させ、生物進化を多様化、複雑化への方向へ進める原動力になった。すでに光合成で色素を使い、鞭毛など細胞運動のメカニズムが存在する生物界で、これに陽イオンチャンネルがリンクするのは当然だろうが、この結果が生物に与えた影響は想像以上に大きい。
例えば視覚を考えると、この重要性がよくわかる。人間の存在しない地球では、光刺激の起源はほとんどが地球から遠く離れた太陽や月で、ここから発する光は生物が生きる環境からの刺激として最も重要なもので、実際多くの生物が光に支配されて活動している。この光の変化に合わせて活動しやすい様、環境をゲノムへと自己化した慨日周期をほとんどの生物が持っていることはこの証拠と言っていいだろう。しかし、この光の元は遥か離れた空の上の、生物が到達できない場所にある。すなわち、視覚は私たちの周りの環境の範囲を急激に拡大させた。
物理刺激のもう一つの特徴は、変化が早いことだ。太陽の光は持続的でも、あっという間に遮ることができる。光のスペクトル(色)の変化になると、反射する物体が変化することで刻々変化する。
このように物理的刺激の受容が可能になることで、環境として生物が受容できる領域は無限の空間範囲へと拡大し、また変化のスピードも速くなった。この結果、(認識するしないに関わらず)生物は空間的・時間的に膨大な環境の変化によく言えばアクセスできるようになり、悪く言えばさらされ続けることになった。
この結果、生物は膨大な物理的刺激に関する情報が溢れる中で生活することを運命づけられる。これに対し、味覚や嗅覚は広がりがない。この差は、感覚が脳のマッピング能力と結合した時大きな違いとして現れるが、これは後で議論する。
脳の発達:トップダウンの神経細胞形成
以前に(http://www.brh.co.jp/communication/shinka/2016/post_000021.html)述べた様に、神経系が特定できる最も下等な動物はイソギンチャクなどの刺胞動物や、クシクラゲに代表される有櫛動物だが、これらに認められる神経細胞は、階層性のない分散型ネットワークを形成している(図1トップ)。このような動物では神経細胞は腹側の上皮層でバラバラに分化し、身体中に広がりネットワークを形成する。系統上もう少し進んだアメフラシになると神経系の階層化が始まり、神経細胞が集まった神経節がうまれ、そこから伸びる神経軸索は束ねられる。神経節では、様々なインプット、アウトプットを連結することで、ネットワークを複雑化することが容易になる。そしてこの究極に、神経系の全てと結合した脳が現れる。
図1:神経構造の系統樹: Linda Z Holland et alの論文より転載(Hollad et al, EvoDevo 4:27, 2013)
こう述べると、神経節や脳の進化が、ポリプのように分散して存在する神経細胞をまず神経節へとまとめ、最後に神経系全体が結合し合う脳へとまとめていく、ボトムアップの過程を想像してしまう。しかし実際には、分散型の神経ネットワークと、脳を持つ神経系の発生様式は根本的に異なっている。
脳を持つ動物の代表は、昆虫と、脊椎動物で、図1からわかる様に、昆虫に代表される旧口動物と脊椎動物に代表される新口動物は、それぞれ独立して進化したと考えられている。にもかかわらず、脳、神経節、神経束、そして末梢神経と続く神経系の階層構造はよく似ている。しかも、この類似性は構造だけではなく、それぞれの神経領域を決定する遺伝子群の発現にも見ることができる(図2)。
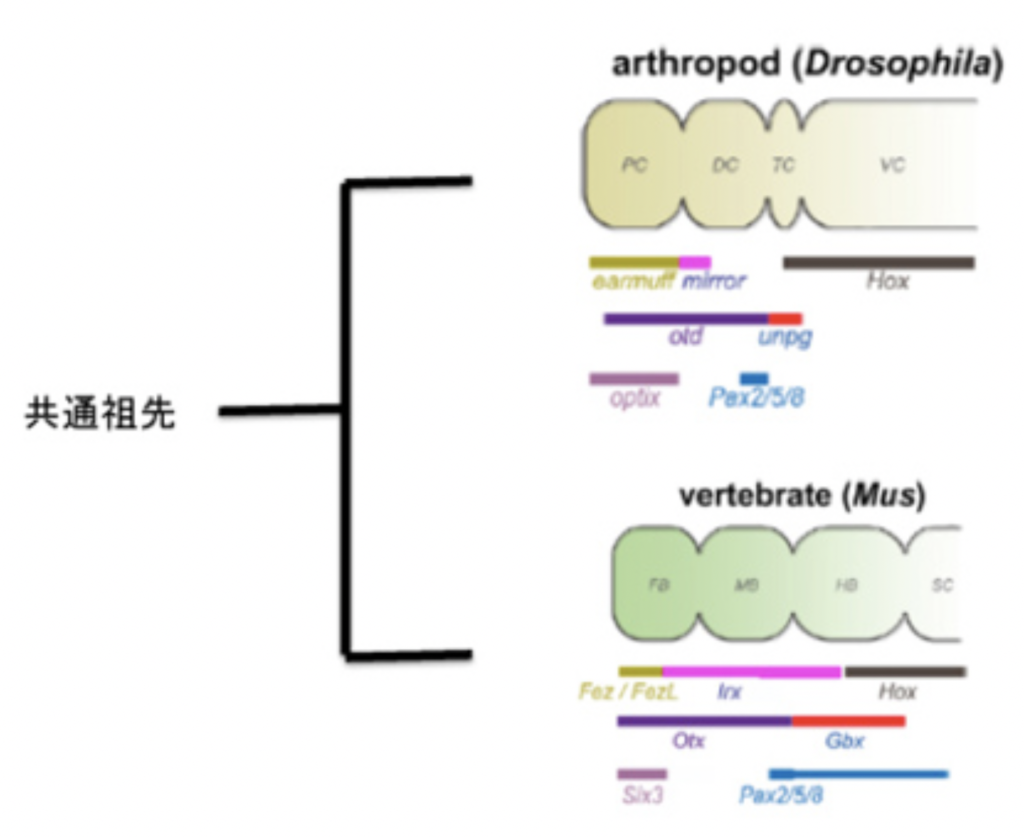
図2 ショウジョウバエとマウスの脳の領域化に関わる遺伝子:Hollandの論文より転載。同じ色で示した遺伝子は、相同遺伝子。
これら階層化された神経系の発生では、ポリプのように神経細胞が個別に上皮から分化することはない。昆虫についてはショウジョウバエしか知識はないが、神経はすでに領域化されたそれぞれの体節で上皮から分化した神経芽細胞が、広がることなく局所で分裂し神経細胞塊をまず形成する。脳は、体節のない頭部に神経芽細胞の集団が現れ、分裂して将来脳になる塊を形成する。
すでに領域化した上皮細胞から神経細胞が形成されるのは脊髄動物も同じだ。脊椎動物の場合、上皮からまず神経管が形成され、神経管の中に神経細胞が分化増殖して細胞集団を形成する。また、頭部も同じ様に、領域化した上皮が陥入し、それが神経細胞へと分化することで形成される。このように昆虫も脊椎動物も、神経が分化する前にボディープランに合わせた上皮の領域化が起こり、領域化された細胞が神経細胞へと分化することで領域性と階層性をうまく両立させたトップダウン方式の神経発生が行われる。この方式は、体節を基礎とする分節化したボディープラン形成という新しい発生過程の進化にあわせて新たに生まれた神経発生様式といっていいだろう。
新口動物と旧口動物は独自に進化したと考えられているが、両者に見られる神経系の類似性から、両者の分岐点に、神経発生をボトムアップ型からトップダウン型へとシフトさせた共通祖先がいるのではと考える人が多い(図2)。
これに対し、ギボシムシに代表される、脳を持たない半索動物(図2、3列目)も新口動物に分類されることから、この類似性は偶然の産物で、脳のような究極の階層性を実現するためには、まず上皮の領域化として現れたボディープランの助けを借りる以外の方法はなかったからだと考える人たちもいる。
いずれにせよ、脳の誕生により、脳に全てを集中し、また脳から全てが出ていくという階層を誕生させることが、神経系が新しい機能を続々と開発した最も大きな基礎となった。
表象とマッピング
脳という神経ネットワークの集中が可能にした最も偉大な機能は、身体や環境の変化を神経ネットワーク情報に転換し(表象し)、対象の空間や時間関係をマッピングする機能だと言える。少し抽象的でわかりにくいと思うので、具体的例を見ながら説明しよう。
まず図3を見てもらおう。これは、私たちの体各部位の運動・感覚神経が、脳のどの部分と結合しているかをマッピングしたもので、ペンフィールドのホムンクルスとしてよく知られている図だ。この図は最初、カナダの脳外科医ペンフィールドが、意識を保ったままの開頭手術の際に、脳を刺激して体の各部位と脳の領域の関連を記録した研究に基づいて描かれた。大脳皮質の運動野と、感覚野にほぼ同じ様な小人の図を書くことができる。
図3 この図は、例えば体の各部位の感覚が脳に集められており、各部位が投射する場所が大脳皮質上に地図として描けること、言い換えると大脳皮質の感覚野に表象されていることを示している。
末梢と中枢の神経投射が脳皮質上に一定のパターンで分布するのは当然だと考える人は多いはずだ。しかし、この図は決して末梢との連結だけを反映しているのではない。末梢から刺激が繰り返されると、今度は脳自体の回路に体の各部からの刺激が記憶され、神経連絡がなくなってもこのパターンを保存することができる。すなわち脳回路の特性へと転換された表象は、末梢からの刺激が途絶えても、末梢の感覚として維持される。このことは、何らかの原因で急に腕が切断された人たちが、あたかもまだ腕がある様に錯覚する幻肢という現象を経験することからわかる。この現象は当初、切断された断端に残った神経から直接脳へ信号が送られるため生じると考えられていた。しかしその後の研究で、私たちの脳内に形成された腕の各部に対応する脳内の刺激の記憶が、末梢からの信号が途絶えた後も、自発的な興奮を続けることで、錯覚を発生させると考えられる様になっている。すなわち、末梢と直接的連結が失われても腕の記憶を呼び起こせるということは、脳自体の神経ネットワークの特性として、腕が表象できていることを示している。
幻肢では、私たち自身の体が脳に表象されるが、脳が表象できるのは体にとどまらず、私たちの周りの環境も同じように表象されていることを明確に示したのは、2014年のノーベル賞に輝いたオキーフ及びモザー夫妻による、場所細胞とグリッド細胞の発見だろう。図4はノーベル賞受賞理由に使われた図1、2を和訳してそのまま転載したものだが、私たちの体と物理的結合や関係のない場所情報が、脳内の場所細胞やグリッド細胞の空間的配置として表象されていることを示した。
図4:場所細胞とグリッド細胞 (C) The Nobel Assembly at
Karolinska Institute
このように脳が誕生したことで、様々なインプットやアウトプットを脳内の神経ネットワークのパターンとして表象することが初めて可能になった。
ここで大事なことは、環境の空間的時間的変化を、脳内の神経回路の特性として表象することができるということは、まさに以前紹介したチャールズ・サンダース・パースのシンボル記号関係を環境や身体と脳回路の間で形成することができるようになっている点だ(http://www.brh.co.jp/communication/shinka/2016/post_000012.html)。
このマッピング能力、表象能力に、記憶が合わさると、自己や意識が始まるが、これについては次回に回す。
[ 西川 伸一 ]
脳の自己
2017年2月15日
脳を頂点とする階層的な神経回路による新しい情報システムの誕生で可能になったことについて書いた前回の内容をまとめると、
になる。
古来人間の最大関心事である自己や意識はこれを基盤として新たに生まれた高次情報だが、その発生のためには、脊椎動物の脳構造の大きな変化が必要だったので、まず構造の進化を簡単に確認しておこう。
脳の急速な増大
脳神経回路を媒体とする新しい情報システムの進化過程では、一貫して脳の神経細胞数が増え続けてきた。例えばマウスは約2億個のニューロンがあるが、人間ではその数は1600億個にのぼる。闇雲に回路を複雑にすれば高次機能が生まれるわけではないが、素子数を増やすことで発展してきたコンピュータを見れば、まず細胞数を増やすことが必要であることは直感できる。実際全く異なる情報システムであるゲノムも進化の過程で塩基数が増大し、ゲノム内の構造も複雑化してきた。
図1:進化の過程で前脳が急速に拡大する。これに続いて、サルやヒトでは皮質を陥入させ脳のシワ(脳溝)を形成することで、体積以上の皮質の拡大が可能になった
脳は大きく分けて、後脳、中脳、前脳、および嗅球に分けることができるが、図に示したように脊椎動物進化では、もっぱら前脳を拡大させてきた(図1)。
前脳皮質の神経細胞を増加させるメカニズムだが、脳室のsubventricular zone (SVZ)で水平に幹細胞を未分化なまま増殖させて皮質面積を拡大するとともに、増殖している幹細胞が今度はradial glial cellに沿って縦に細胞を供給し、順番に異なるタイプの神経細胞へと分化することで縦に厚みが増すことで達成されている(図2 Montiel et al, Journal of Comparative Neurology, 524:630, 2016 より転載)。
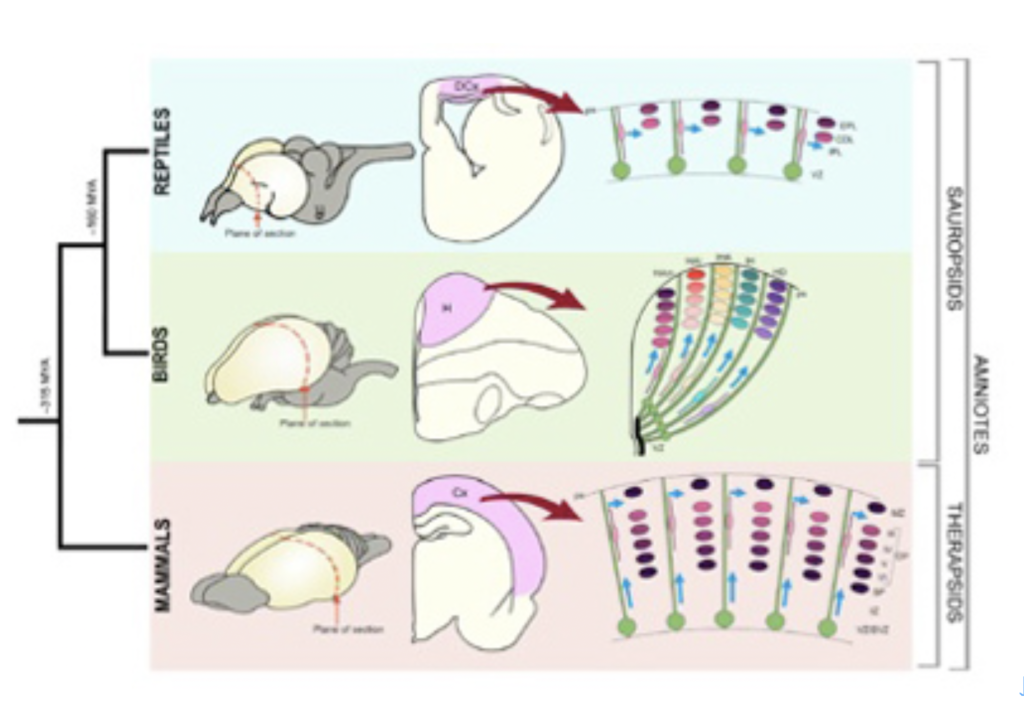
図2 哺乳動物、鳥類の皮質神経細胞を縦に並べていく神経形成の様式について示している。神経細胞を作る幹細胞がまず水平に広がり、この細胞から造られた神経細胞はradial
glial cellに沿って移動しながら分化し、細胞数を増やす。
ただ脳の表面をそのまま増大させるだけでは、脳のサイズという物理的限界に当たる。そこで脳進化の過程で、皮質が立体的に陥入するいわゆる脳のシワ(脳溝)が形成される様になり、このおかげで、サルから人間の進化の過程で脳の皮質面積は急速に増大する(脳溝は図1の人間の脳参照)。
同じ幹細胞に由来する異なるタイプの神経細胞が縦に並ぶことで脳皮質の層構造(6層)ができる。この縦に並んだ細胞からできる層構造が最小セットとなり、この単位が横に束ねられ、細胞同士がネットワークを作ると、カラムと呼ばれる脳の最小機能単位が出来上がる。このカラム構造では、各層の神経細胞の反応性は似通っている(例えば同じ方向の動きに反応するなど)。この様に、脳では細胞数がただ増えるのではなく、小さな単位のネットワークが集まってモジュールができ、それがまた集まってという様に、ネットワークは階層的に拡大していく。
以上が脳のサイズ拡大を支える解剖学・細胞学的の知識の基本だ。この基盤の上に、自己や意識が生まれる。
自己の誕生
自分の行動を他の動物と比べ、また自分の過去を振り返れば、自己や意識が脳の進化とともに現れ、また「物心つく」と言うように、自己や意識が一人一人生まれてから新たに発生することがわかる。もちろん古来、自己や意識は人類にとっての最大の謎で、哲学から生物科学、神経生物学、心理学まで多岐にわたる議論が続けられてきた。当然この話題についてのすぐれた著作は多く、本当は私の出る幕ではない。しかし脳神経回路を媒体にする情報の理解なしに、生物の情報の進化を構想することはできないので、この問題についても私なりの理解をまとめておくことにした。
まず問題は、自己から始めるか、意識から始めるか、あるいは同時に扱うかだが、両者は脳の最も高次の機能として密接に関連し合っているため、どちらがより重要とか、どちらが高次とか決めることはできない。
脳神経系以外の自己
そこで「エイヤ!」と「自己の誕生」から始めることにした。というのも、自己という言葉は、脳神経回路情報だけでなく、様々なレベルの生物活動に対して使われる。私自身も原始ゲノム誕生について説明した時、「ゲノムの自己性」と、この言葉を用いた。実際、私たちは「細胞の自己」のように脳神経系が存在しない生物に自己を使っている。例えば自己複製という言葉は最もポピュラーだろう。他にも、昨年のノーベル賞に輝いた大隅良典さんの「オートファジー」も、細胞の自己を想定した使い方だ。
これらの場合で、「自己」の意味する内容を考えてみよう。ゲノムの自己と私が呼んだ場合はひと塊りの核酸が周りから完全に分離し、その上で自律的な複製能力を持つことが条件となっている。これが単細胞動物となると、内と外を隔てる細胞膜が存在するため周りからの分離はさらに明確になる。ただ、シャボン玉のように膜で周りから分離される状態が維持できても、自己性があるとは思わない。周りの環境から完全に分離するとともに(自己と外とに境界線を引くことができる)、同じ細胞(境界の内側)を自律的に複製できる能力があって初めて自己性があると言える (熱力学的条件などは自己複製で全て表現できている)。周りから完全に分離して、自己複製能を持つことが、「自己性」成立の条件だ。
多細胞動物になっても、個体の内と外ははっきりと分けることができるため、細胞と同じ意味で「個体の自己」を使うことができる。ただ場合によって「個体の自己」と「細胞の自己」が分離することがある。最も分かりやすいのがガン細胞で、個体から独立して勝手に増殖すると同時に、ゲノムも個体とは異なっていると言える。この極端な例が、ハマグリやタスマニア・デビルに見られる、個体から個体へと伝搬するガン細胞だろう。このようなガン細胞は自己性が高いと言えるかもしれない。
他に「自己」が定義されている分野が免疫学だ。
免疫学はジェンナーによる種痘の成功により認識された「2度なし現象」(一度感染すると次に感染しない)についての研究がはじまりで、パストゥールの狂犬病ワクチン(抗原の概念)、そして北里・ベーリングによる免疫血清療法(抗体の概念)の開発により、外来抗原に対する特異的な抗体反応という概念が確立する。
このように最初、外部から侵入する病原体への防御反応機構として始まった免疫学は、ランドシュタイナーらによる血液型の発見により、免疫系が自己の成分と非自己の成分を区別できることが明らかになる。その後、臓器や組織の移植研究から、移植抗原が(MHC)が異なる他人からの組織に対してだけ強い免疫反応が誘導されることが明らかになると、「自己と非自己の区別」は免疫学の最重要課題になる。
20世紀後半に始まる免疫学の大きな進展により、免疫系が自己成分とそれ以外をどう区別するかについてはすでに詳しくわかっている。詳細を省いてこのメカニズムを見てみよう。
図3:免疫系の自己と非自己の識別:説明文中
免疫反応にはT細胞とB細胞が関わっているが、原則的にはどちらの細胞も自己の成分に対しては反応しない。これを、自己成分に対する免疫寛容と称している。この寛容には2種類の戦略が存在している。最初の戦略は、リンパ球の分化がまだ未熟な段階で抗原刺激を受けると、成熟後におこる細胞増殖や分化の代わりに細胞死が誘導される。発生初期から体内に存在する自己抗原は、未熟な段階のリンパ球と反応することができこの結果自己の抗原に反応するリンパ球の細胞死を誘導して免疫システムから除去している(図3)。この過程は、T細胞では胸腺内、B細胞は骨髄内で行われる。
もう一つは我が国の坂口志文らにより明らかにされた戦略で、細胞死による寛容とは逆の戦略と言える(図3)。この戦略は制御性T細胞と呼ばれる特別なポピュレーションだけで見られる。制御性T細胞が自己抗原と出会うと、細胞死の代わりに活性化され、長期間体内で生存できるようになる。これにより後にキラーT細胞のような他のリンパ球集団が同じ自己抗原に反応しようとするとき、その細胞を抑制し、自己抗原に対する反応を抑制する。
このように免疫系の多様な認識システムは、発生過程で生成的に形成される。こうして生成される免疫系の自己とは、免疫系の抗原受容体が認識できる自己抗原の総体と定義でき、これらの抗原は免疫系が反応できないレパートリーとして免疫系に表象される。言い換えると、自己抗原は発生初期の選択の結果、細胞死による「レパートリー欠損」として、あるいは自己反応性抑制性T細胞レパートリーの長期維持として表象されている。自己の範囲に境界線を引くとすると、免疫の自己とは、成熟後の免疫系が反応できない抗原として線引きができる。
実は免疫システムにはもう一つの自己認識がある。それはMHC restriction(組織適合抗原拘束性)と呼ばれるT細胞の認識方法だ。抗体と違ってT細胞の抗原受容体は、抗原をそのまま認識することまずない。代わりに、図4に示すように、まずタンパク質はプロテアソーム(タンパク質分解酵素複合体)でペプチドへと分解され、それが自己のMHCと結合したときに新たに生まれる分子構造を認識している。
図4 T細胞抗原受容体は、一度自己MHCに参照できたペプチドだけを認識する。Wikipediaを改変転載。
言い換えると、自己、非自己を問わず、T細胞抗原受容体に対する抗原は小さなペプチド+自己MHCで、抗原は自己のMHCと結合するかふるいにかけられた後、T細胞抗原受容体で認識できるようになる。すなわち、抗原の段階でまず自己に参照される。ただこの場合も、免疫側にとって自己ペプチド+自己MHCを含む、T細胞抗原受容体が反応できない全ての抗原が自己と認識され、これは抗原に反応できるT細胞受容体のレパートリー中にぽっかりと空いた欠損として表象される。
以上、脳神経系の自己に行き着く前に、脳神経系とは異なる2種類の自己の定義について見てきた。この2つの自己をまとめておくと、
の2種類があることがわかってもらえたと思う。
一見これらは、脳神経系の自己とは接点がないように思えるかもしれないが、脳神経系に生まれる自己性は、この2種類の自己と多くの共通性を持っている。次回はいよいよ、脳神経系に生まれた自己について見てみよう。
[ 西川 伸一 ]
脳神経系の自己基盤と原始自己
2017年3月1日
個体が生まれた時自己としてのゲノムは既に決まっており、生きている間ほぼ変わることはない。しかし免疫系の自己のように、生まれたときから決まっていないものもある。前回説明したように、免疫システムの自己は、発生・発達過程で、外界、内界からの刺激に応じて生成的に形成される。
免疫系の自己と同じで、脳神経系に生まれる自己も、脳の機能がほぼ完成した後、体の内側、外側から受け続ける脳神経系への入力(経験)の結果として生成的に形成される。注意しておきたいのは、免疫系の自己とは異なり、脳神経系では、脳が受容できる入力の情報量、表象能力、情報の連合能力などは動物ごとに大きく異なっている点で、この結果、入力に応じた脳の受動的変化が積み重なった原始的な自己から、後に議論する意識も関わる能動的に形成される高次の自己まで、自己に大きな多様性が存在する。人間の脳神経系の自己はこの全てのレベルの自己が統合されて形成されているが、最初はまず内外からの入力の影響が積み重なって受動的に生成する最も原始的な自己の形成から考えてみよう。
まず脳神経系の自己を発生させるためには、それを可能にする脳の構造・機能的基盤の発生が必要になる。この脳神経系の構築に関わる最も重要な情報がゲノムで、進化、あるいは種内の個体間の多様性として生まれるゲノム情報の違いは、脳神経系の構造や機能の違いに反映される。
例えば、魚から人間まで、脊椎動物はゲノムの違い(進化の結果)に応じて、構造的・機能的に異なる脳を持っている。当然脳神経系に発生する自己にはこの構造的、機能的違いが反映される。ゲノム進化で生まれる大きな違いほどではないが、個体間に小さなゲノム変化が生じただけで脳機能の異常が生じ、様々なレベルの自己の発達障害が起こることが、人間のゲノム解析や、遺伝子操作動物を用いた研究から明らかになっている。とはいえ、自己や意識の形成に関わるゲノム進化に関するリストが増えても、構造や機能と対応付けて説明することは今なお難しい。
さて、脳神経の自己が生成的に形成されるということは、イギリス経験論を説明するのに例えとして使われる、「人生は何も書いてない白紙から始まる」ことを認めることだ。この白紙が入力を経験したことがない生まれたばかりの脳で、この白紙に刻々かわる経験の表象が記録されることで、独自の自己が形成されると考えればいい。
これまで見てきたように、脳を持つ神経系は、
を持っている。
これらの機能のおかげで、もし全く同じ構造の脳神経系が発生したとしても、経験する入力が違えば、それによって誘導される脳の回路の特性も異なることになる。このように、最も原始的な脳神経系の自己とは、ゲノム情報に強く支配された発生過程で形成された基盤(「自己基盤」)が、異なる経験を重ね、その一部を記憶することで、独自の構造的・機能的特性を持つ脳神経系へと変化することだと言える。
この意味で、アメフラシのような単純な脳神経系の自己基盤も、刺激を神経興奮に転換して表象し記憶することができることから、原始的自己を持っていると言える。経験したエラの刺激の強さや回数で、水管反応の特性が変わるが、この結果を我々が外から観察する時、アメフラシの間に個性が生まれると考える。これをアメフラシの側から見ると、他の個体とは異なる自己ができたことになる。もちろん、アメフラシは自己が誕生したと認識はできていないが、この点については次回以降に考察したいと思っている。
このように経験と記憶を重ねることで積み重なる神経回路の変化が原始的自己形成と考えることができる。実際以前紹介したように(http://www.brh.co.jp/communication/shinka/2016/post_000024.html)、記憶過程は神経細胞のシグナル伝達経路や、遺伝子発現を変化させ、神経伝達因子に対する反応特性を変化させるとともに、シナプスの形態学的変化を誘導して、神経の結合性の長期変化を誘導できることがエリック・カンデルたちにより示された。もちろん、哺乳動物の脳のようにはるかに高次な脳神経系でも基本的に同じメカニズムを共有している。
最近になって神経同士の結合を支えるシナプス構造のダイナミックな変化を直接観察する研究が進んでいる。この分野の研究を知ると、私たちの脳でいかに膨大な変化が刻々起こっているのかを実感できるので、すこしだけ最近の論文を紹介する。
哺乳動物の脳でのシナプス形成の中心は、神経細胞の樹状突起から飛び出たスパインと呼ばれる構造と神経軸索との接合により担われている(図1)。この時、刺激は必ず神経軸索からスパインへと伝達される。
図1:スパインの構造:左写真に示すように、スパインは様々なサイズ、形態を呈する、神経細胞樹状突起から飛び出た突起だ。これは神経軸索と結合して、軸索の興奮を受け取る役割がある。右図に示すように、スパインは様々な形態をとるが、この形態の違いがシナプス伝達の特性の違いに結びついている。(出典:Wikipedia)
同じくウィキペディアから転載した図2を使ってスパインのダイナミズムを説明しよう。図2のAからEは一本の樹状突起から出るスパインの変遷が示されているが、樹状突起はスパインを伸ばして、複数の神経軸索と結合し、興奮を受け取ることができる。図に示されているように、学習によりスパインと軸索の結合はダイナミックに変化し、回路の特性が変化すると考えられてきたが、スパインの消長を実際に観察することは難しかった。
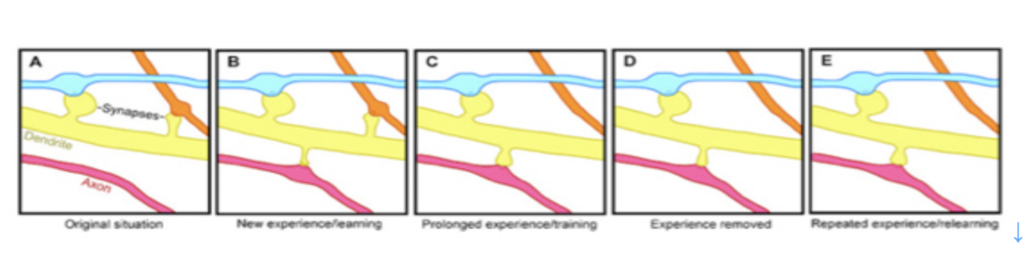
図2:学習過程でのスパインの変化(出典:Wikipedia)
ところが2015年夏スタンフォード大学から、生きたマウスの脳内の樹状突起から飛び出すスパインをなんと22日間にもわたって観察し続けた論文が発表された(Attardo et al, Nature, 523:592-596, 2015:論文はウェッブからアクセスできる:https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4648621/)。
この研究では、樹状突起の細胞膜が蛍光を発するように操作したマウスの脳に一種の内視鏡のような長焦点レンズを挿入して、樹状突起のスパインの消長を22日間観察している。実際の像については論文を参照してほしいが、図2で示したよりはるかにダイナミックなスパインの消長が起こっている。また、中には極めて安定なシナプスを形成しているスパインが存在することもわかる。
同じ方法を用いて、学習から睡眠中という短い時間サイクルで大脳皮質のスパインがダイナミックに変化することを示した論文も最近発表された。(Ma et al, Nature Neuroscience, doi:10.1038/nn.4479)。この研究から、学習時によって一本の樹状突起から軸索に伸びるスパインの数は増加するが、睡眠(REM睡眠:睡眠中も盛んに目が動いており、夢を見ている時はこの睡眠時期にある。覚醒中に学習した記憶を整理して必要な記憶を固定化する過程に関わると考えられている)に入るとスパインは剪定され数が減る。しかし、一部のスパインではより大きな終末をもつスパインへと構造変化が起こり、このようなシナプスは長く維持されることが明らかにされた。
これはマウスの大脳皮質での観察だが、一回覚醒と眠りのサイクルを繰り返すうちに、一本の樹状突起のスパインに、目に見えない生化学的変化だけでなく、シナプスの数や構造といった目に見える形態的な変化が刻々起こっていることが明らかになった。スパインの数は1000兆を越すと考えられているが、毎日これらのスパインが発生、剪定を繰り返し、その中から長期間安定なシナプスが形成されている。このダイナミックな過程を直接観察できるとは、今後の解析が楽しみだ。
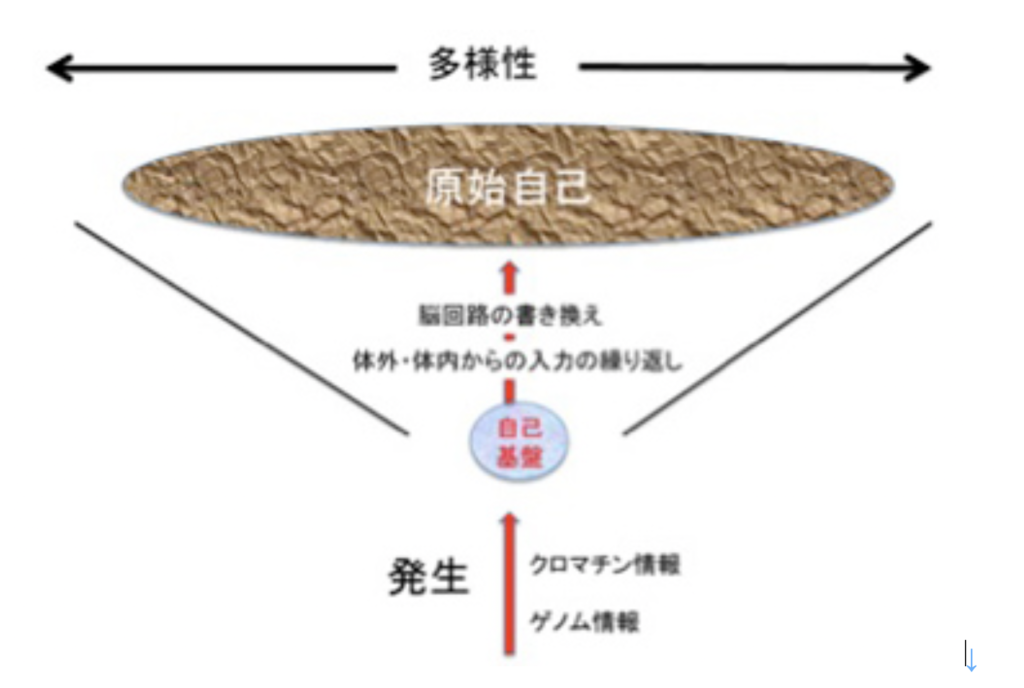
図3:自己基盤から原始自己へ:自己基盤はゲノム情報に従う発生過程により生まれる。この自己基盤としての脳回路は、体内外からの入力により書き換えられ続ける。
脳を持つ生物が生きている間様々な経験を繰り返すと、その入力により脳回路は不断に作り直される。この結果経験が異なれば、まったく同じ脳構造から始めても、それぞれの個体の脳構造は自ずと違ってくる。すなわち、独自の個性が発生する。重要なことは、この違いを生み出す原理は単純で、入力により神経回路を構成する神経細胞のスパインの数や構造を変化させておこる神経細胞同士の結合の強さの違いが生まれるだけだ。しかし、脳の細胞数が増大するにつれて、異なる強さで結合した神経同士から形成される回路は、ほぼ無限の多様性を持つことができ、無限の個性、無限の原始的自己が発生する。
私の勝手な定義と断った上で、以上をまとめると、脳神経系の原始的自己とは、生物個体の脳神経の個性のことで、経験入力により回路が書き換えられた後の脳神経系だ。従って、脳神経の原始的自己も刻々変化する(図3)。重要なことは、この脳神経系が経験する入力は必ずしも個体外からの刺激だけではなく、体の表面、内部からの刺激も入力として扱われ、原始自己の形成に関わっている。これは、免疫の自己も同じと言える。ただ私たち人間の脳と比べると、こうしてできた原始的自己は、自分で自己として認識されることはない。すなわち、脳神経システムが、脳神経システム内の自己を認識できるようになるためには大きな転換が必要になる。これについては次回以降考えることにする。
その代わり、最後に免疫系と脳神経系を比べて終わりたい。免疫系も、ほぼ無限と言っていい抗原に対して反応することができ、自己と他を区別できる。しかし、神経系と比べるとその複雑性には限界がある。この原因として、脳神経系と免疫系の情報を扱う形式が異なっていることがあげられる。
図3で示した脳神経系の原始自己と比べた時、免疫系では抗原(T細胞抗原としてのペプチドも含めて)は、抗原受容体を刺激し、刺激された細胞の特異的な増殖、生存、細胞死を誘導して、免疫担当細胞のレパートリーを書き換えることで経験が表象され、記憶される(図4)。
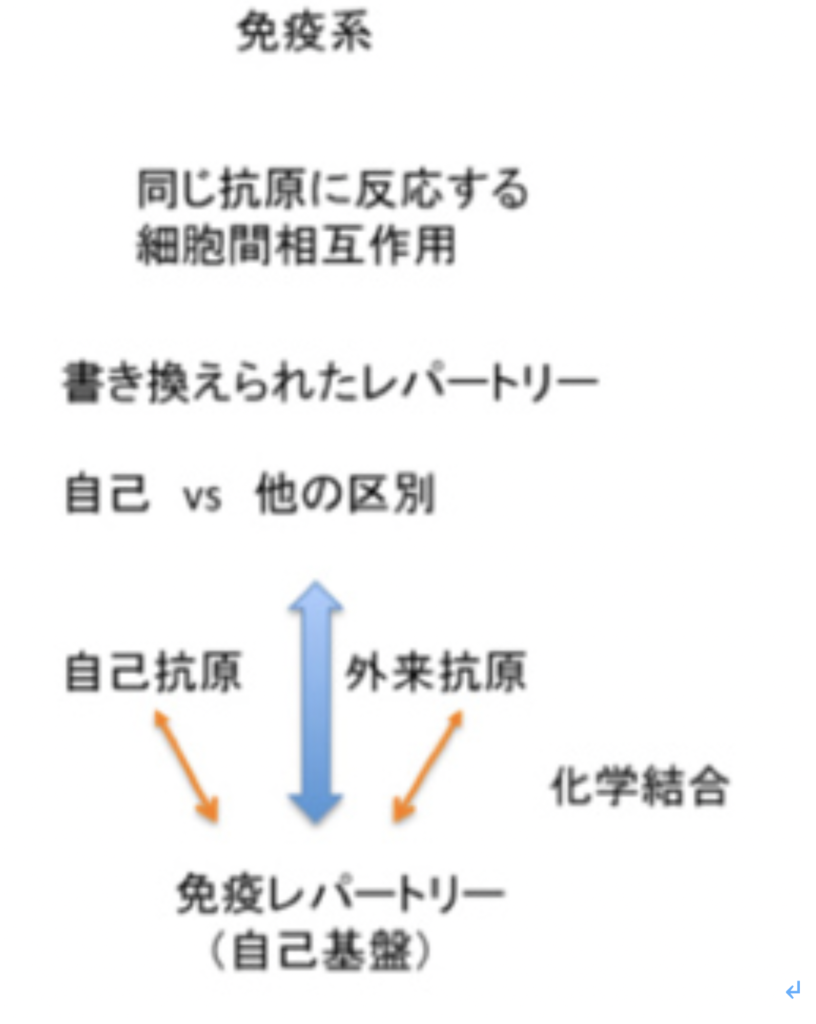
図4 免疫系では外来と自己抗原の区別は、反応性抗原受容体のレパートリーの差として表現されている。
レパートリーと表現したのは、個々の免疫担当細胞が異なる抗原受容体を持っており、この細胞が様々な比で集まったものが免疫系だからだ。抗原と抗原受容体は化学的に結合し、免疫担当細胞のレパートリーも抗原との化学的結合可能性と表現することができる。また、免疫システムの反応も、抗原との化学的結合を必ず前提として行われる(図5)。
ぜひここで思い出していただきたいのは、パースの記号論だ(http://www.brh.co.jp/communication/shinka/2016/post_000011.html)。抗原刺激という経験と、その表象、記憶、反応が、抗原vs抗原受容体との化学結合を基盤として持っている点で、免疫系はパースのいうインデックス記号関係を基盤にした情報システムと言っていい。
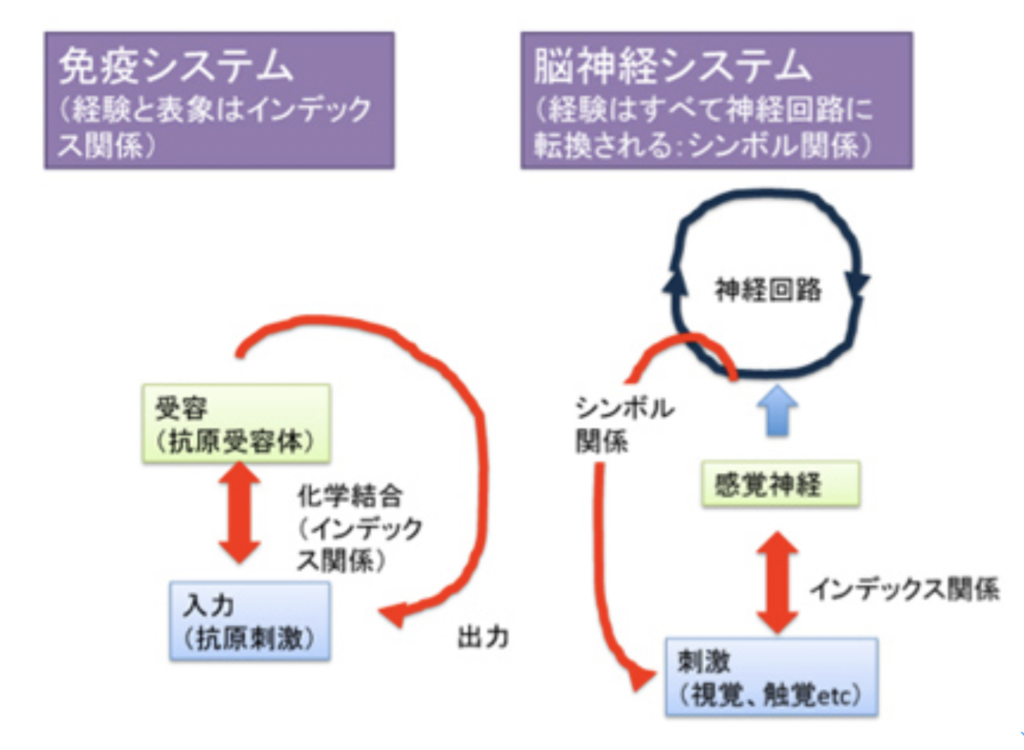
図5:免疫系では経験とシステムの間にインデックス関係が成立しているが、神経系では、刺激はインデックス関係を経て、さらに神経回路とのシンボル関係へと変換される。
これに対し、脳神経系では入力はまず感覚神経の興奮を誘導する。この入力の時点では、感覚細胞内で光子が色素を活性化したり、ニオイ物質が嗅覚受容体に結合することで神経が興奮するため、ここでもインデックス関係が成立していると言える(図5)。しかしインデックス関係が成立するのは感覚細胞内までの話で、感覚が神経膜の興奮に転換され、全く別の神経回路として表象されると、経験と、それに対応する神経回路との間に物理・化学関係は完全に消失している。この意味で、脳神経系の経験(刺激)とその表象と記憶の関係は、パースのシンボル関係と言える。
脳神経回路を基盤とする情報システムは、おそらくDNA情報に続いて地球上に誕生した新たなシンボル情報の誕生と言っていいだろう。このシンボル情報であるということが、ほぼ無限の複雑性を獲得する重要な契機になったが、そのためにはまず意識と、脳神経回路の自己を認識できる「脳神経回路」の誕生が必要だった。
[ 西川 伸一 ]
意識と自己
2017年3月15日
ベッリーニのオペラに「夢遊病の女」という不思議なタイトルの作品がある。結婚前夜に夢遊病のために無意識になった主人公が伯爵の部屋に迷い込んで寝込んでしまうが、他の男のベッドで寝ているのを知った婚約者が「不倫だ、破談だ」と大騒ぎになる。無実の主人公は悲嘆にくれるが、今度は村人が集まっているところに、夢遊病発作で再び無意識に陥った主人公が現れ、無実が証明されるという、荒唐無稽な話だ。 音楽は美しいのだが、実際の舞台を見るといつも違和感が残る。ベッリーニがどんな意図でわざわざこのテーマを取り上げたのか不思議に思っていた。
ところがシチリア、カターニャの大聖堂を訪れた時、ベッリーニがこのテーマに並々ならぬ意欲を持っていたことがよくわかった。大聖堂には彼の棺が祀られているが、なんとこの墓のレリーフには、最もよく演奏される「清教徒」や「ノルマ」を差し置いて、夢遊病の女のシーンが、音楽の一節とともに使われていた(図1)。

図1 カターニャの大聖堂に祀られている、ベッリーニの棺の台座に描かれたレリーフ。オペラ「夢遊病の女」の一場面で、夢遊病発作に陥ったアミーナを見て村人たちが驚いている場面と想像している。
しかし本当に夢遊病はあるのだろうか?卒後7年ほど医師として過ごしたとは言え、内科だった私には実際の患者さんを見た経験はないし、心の片隅ではまだ信じられない。しかし数多くの論文が毎年発表されおり、総説を読んで見ると、ベッドから起き出して歩き回り、そのことを覚えていないという症状はなんと15%以上の子供に見られるらしい。
眠りとの関係も明らかになっており、年齢を問わず夢遊病で徘徊するときは、ノンレム睡眠と呼ばれる最も深い睡眠時に入っていることがわかっている。すなわちゆっくりした脳波を示す熟睡状態のまま、患者は歩き回っている。驚くことに、服を着替えて車を運転したという報告もある。徘徊中は図1に示したレリーフからわかる様に、目は大きく見開き、背筋を伸ばして歩いており、障害物も的確に避けることができる。しかし、呼びかけても気がつかないし、普通は眠りが覚めたら徘徊中のことはほとんど覚えていない(総説によると(Blake and Logothetis, Visual Competition, Nature Review Neuroscience, 3:1 (2002))実際には6割程度の患者さんが発作中の経験を一部ではあるが思い出せるらしい)。
その存在を信じる、信じないは別として、夢遊病は、私たちが通常「意識」と呼んでいる機能が喪失しても、外界を感知しそれに合わせた行動を取ることが可能であることを示している。言い換えると、外界の知覚、その表象、記憶を呼び起し、イメージの統合、行動の決定から実行までが、「意識」なしに可能であることを示している。
夢遊病の例を最初に持ち出したのは、もちろん今回から意識の問題を取り上げて、脳神経系の「自己」の問題と一体化して考えていこうと思っているからだ。ただ意識とは何かと大上段に振りかぶって考えるのは、私の任ではない。古来、意識については様々な説明がなされ、現在なおその本態については議論が続いている。
他の脳機能の単純な延長に過ぎず、わざわざ「意識」として抜き出して考える必要はないという意見から、神経系を持つ動物全てに存在するという意見、あるいは人間だけが意識を持つとする説まで様々な考えがある。このため、以後の議論は、意識の定義には深入りしないで、私たちが普通、覚醒時に行動するときには存在して、夢遊病の発作時の行動には存在しないものを意識と、まず漠然と定義しておいて進める。
もちろん熟睡時や、麻酔で眠っている時も意識は存在しない。しかし夢遊病の例は、意識がなくとも私たちは、リアルタイムに入ってくる感覚やこれまでの経験に基づいて行動できることを教えている。しかし意識がないと、新しいことを計画して行動することはない。このように意識の存在はこれまで経験したことのない新しい認識を可能にし、将来に向けた計画的行動には必須の条件になる。
睡眠中や麻酔中に意識は失われるが、意識は覚醒していることではない。覚醒していても、意識せずに行動することはいくらでもある。
私は毎朝ノルディックウォークを日課にしている。ストックを持ってクロスカントリーと同じ要領で歩くウォーキングだ。若い人ならすぐできるのだろうが、私の年になると、最初は自動的に足と腕の動きが揃うことはない。「意識」を集中して、「右、左」とうまく揃うように努力する。しかし100mも歩くと、もう意識しないで腕と足は互い違いに動くようになる。逆に、急に足が止まった後歩き出す時、どの腕を出すのかなどと意識しだすと、逆にうまくいかなくなる。意識を排除して動けるようになると、今度は意識がその動きを邪魔する。
あらゆるスポーツで同じことが見られる。スポーツの練習とは、意識しなくても自然に体が動くようになるまで動作を繰り返すことだ。テニスでも、サーブを打つ相手の微妙な動きを情報としてそれに合わせて走って行く場所を選び、レシーブする一連の動きが意識しないでできるようになると、一人前になる。実際、プロのテニス選手の200km/hを越すサーブがどこに落ちるか見てから行動しても遅い。無意識に動けるからこそ、プロの試合が成立する。どれだけ無意識になれるかが、優秀でクレバーな選手といわれる条件になるのは逆説的だ。
覚醒と意識が全く異なることを一番よく教えてくれるのが盲視と呼ばれる病態だ。何らかの理由でV1と呼ばれる一次視覚野に損傷を受けると、網膜で受けた刺激は知覚できない。しかし、網膜は働いており、そこから直接視床をとおる経路が存在しているおかげで、患者さんは障害物を意識しないまま避けることができる。もちろん患者さんは全く見えないと思っている。当然、景色を眺めて今度はあの道を通ろうなどと新しいことを計画することはない。視覚だけに限れば、夢遊病と同じに見える。しかし、音や臭いについては意識することができているし、明らかに覚醒している。
このように、無意識に個別の運動を支配したり、視覚のみ意識できない盲視の例は、意識が脳内に分散して活動を支配していることを示している。もちろん夢遊病のように意識全体が失われることもあることから、意識を形成する神経ネットワークも階層的なネットワークを持ち、支配する範囲が変えられる様だ。
さてこれまで、意識を夢遊病の発作時に失われる、新しい認識や計画のために必要な何かと定義してきたが、この定義だと意識は人間特有の高次脳機能だという話になりかねない。夢遊病から話を始めたが、私自身、意識は決して高次機能に限るわけではなく、原始的な形は神経回路形成以降かなり初期から存在していたのではないかと思っている。このことを理解してもらうため、意識をもう少し生物学的に調べている研究を見てみよう。
意識の生物学的役割を理解するための面白い例が、両眼視野競争と呼ばれる現象に見られる。両眼視野競争とは、左右の目に異なる像を別々に見せる時、実際にはどちらか片方の像だけが見えて、もう一方が見えない現象を指している。もちろんそれぞれの像は左右の網膜で感知され、反対側の一次視覚野へ投射されている。しかし、実際に私たちが見えていると感じるのはどちらか一方だけで、残りは見えているのに全く認識できない。多くの場合、2つの像が入れ替わり立ち替わり意識に上り、最後はどれか一方の絵が優勢になる。この実験は、私たちは見ていても見えないものがあり、見えるかどうかは意識にかかっていることを示している。すなわち、意識は競合している両眼の像のどちらを認識するかを決めるのに働いているのがわかる。
意識により競合する知覚が選択されるという現象は、なにも両眼競合に限るものではない。図2に示した、見方によって若い女性が見えたり、老婆が見えたりする有名なだまし絵も、意識によりイメージが選ばれていることを示す例だ。

図2 有名なだまし絵:若くて高齢の女性。心理学者エドウィン・ボーリング(Edwin Boring)がこの絵を用いた視覚の研究成果を1930年に発表した。
羽根飾りに目をやると若い淑女が現れ、視線を下に落とすと老婆が現れる。両眼視野競争とは異なるが、だまし絵は意識とは何かを教えてくれる。(出典:wikipedia)
これらの例から、内外からの様々な刺激により脳内に形成される数多くの表象の中から一つの表象を選ぶことが、意識の機能と考えることができる。脳神経回路に入ってくる情報量の膨大さを考えると、この選択システムがないと、脳はすぐにパンクしてしまう様に思う。
もちろん神経系だけが環境からの膨大な情報に曝されているわけではない。安定的に見えるゲノム情報でも、様々な外界からの情報に曝されており、介入を受け続けている。最近注目されているCRISPR/Casシステムも(http://www.brh.co.jp/communication/shinka/2014/post_000009.html)、本来バクテリアが自分のゲノムへの外来ゲノムの侵入を制限するため進化したもので、ゲノム情報が常に外来の情報に曝されており、その情報を制限する必要があることを示している。
また、免疫系はもともと外来の様々な物質に対応する備えとして進化してきた。しかし、これら2つの系では、外来の刺激の認識自体が、例えばDNA同士の組み替え、あるいは抗原と抗体の結合の様に、物理化学的制限を受けている。従って、抗原受容体のレパートリーに認識できる抗原の数は制限される。
一方脳神経システムでは、入り口の刺激情報こそホストの感知能力による制限(例えば音の波長、光の波長などの制限)を受けてはいるが、この程度で情報量を制限することは不可能だ。例えば視覚で言えば、錐体細胞だけでも片方に800万個も存在する。さらに1次視覚野だけで見ても、この情報は刻々かわる。
さらに厄介なのは、光、音、触覚、匂いなど入り口は違っても、結局脳回路に表象されると、入り口の違いに関わらず表象同士で連結することができる。この様に、連結可能性まで考えると、私の脳が例えば1分で晒される情報の量は途方も無い。
このことを電車の中で席に座っている状況で考えてみよう。前に座っている女性の顔はあまり動かないが(ときには笑ったりあくびをしたりしているが)、窓の外の景色は不断に変わり続けている。これに加えて、様々な音が耳に入ってくる。また、足から、お尻から伝わる振動。頭の後ろに感じる太陽の熱。そして、時によって胃がシクシク痛むといった内臓からのシグナルも入ってくる。これを全部総合すると、何ビットになるだろうか。さらに、最初の刺激が異なっていても、神経回路の興奮に還元されると、あらゆるインプットは連結できる。おそらく電車に座って1分で経験する情報は天文学的量にになっているはずだ。このままだと、どんなに神経細胞が増え、回路が複雑化しても頭の中がパンクすることは間違いない。
幸い、ここに意識が登場すると、状況は一変する。例えば前に座った美しい女性に意識を向けると、窓の外の景色も、電車の揺れも、太陽の熱も、胃の痛みすら消えることがある。
逆にどこから来るかわからないが、お腹がシクシク痛むのが気になり始めると、今度は車外の景色や振動どころか、前の女性すらすっかり見えなくなってしまう。
この例から言えるのは、私たちの脳神経系が刻々と曝されている膨大な情報の中から、意識は特定のセットを選び出す役割を果たしているということだ。神経系での情報が神経細胞ネットワークとして表象されていることを認めると、意識が行っているのは、興奮している様々なネットワークの中から少数のネットワークを選んでいることになる。
この様々な神経回路上に形成された表象を選ぶ役割が意識だと定義すると、少し複雑な神経細胞ネットワークを持つ動物は意識が存在してもおかしくはない。たとえば、動物を訓練する時、こちらが期待する行動を取った時だけ、褒美を与えて覚えさせること広く行われている。この時、褒美を与えることで、特定の行動につながるネットワークが選択される。これは、広い意味で意識されたと言える様に思う。
極端な例で言えば、アメフラシにも意識があって何の不思議もない。アメフラシの水管反応を誘導するエラの刺激を繰り返すと、慣れが起こって水管反応が起こらなくなる。そこに電気ショックを頭に与えると、また水管反応が起こるようになる。これは電気ショックを受けた回路の影響で、エラの刺激が再び水管反応の刺激として選択されたことを意味しており、神経回路の選択を意識として定義するなら、最も原始的な意識と呼んでいいと思う。
ここまで、最初は極めて高次な神経活動として意識を提示しておきながら、一転アメフラシの神経系で見られる神経回路の選択まで意識に含めてしまうと、混乱の極みとお叱りを受けそうだ。しかし、何を基準に膨大な情報から選択が行われるのかを考えていくことで、この混乱は収束できる。その点について、最後に少し触れておこう。
両眼視野競合の際に、縦縞と横縞を別々の目に提示したとしよう。ほとんどの人は縦、横に特に好みはないはずで、実際縦縞と横縞が周期的に現れる。おそらく、サルや猫のような両眼で見ている動物では同じことが起こっている。とすると、この異なるイメージが周期的に選ばれるという選択基準は、私たちの脳回路自体にプリセットされていると考えられる。
同じ模様を使っていても、明るさなどが大きく違っている場合は、最終的にはっきりしている方だけが見える様になる。はっきりしたほうを最終的に選ぶ基準も、生まれた時の脳回路にプリセットされていると考えていい。
ところがもう少し複雑なだまし絵になると、どんなに努力しても片方の絵柄が見えないという人も出てくる。この場合、個人の脳回路の特性による場合もあるが、それに加えて個人が過去に得た経験・学習の結果、特定のイメージにバイアスがかかってしまっている可能性もある。
この様に、表象が選択される基準を探っていくと、進化や発生過程で形成される脳回路自体の特性、あるいは学習や経験により新たに書き換えられた脳回路である場合もあることがわかる。また学習による脳回路の書き換えも勘定に入れ出すと、簡単な書き換えから、極めて複雑な書き換えまで多様性が高いと考えられる。この結果、当然同じ刺激により生まれる表象に対しても、選択基準は一人一人、異なっている。
この様に選択する基準について考えてみると、ゲノムから高次の脳回路書き換えまで、極めて多様な基準が働いていること、そしてこれらの低次から高次にわたる基準は、前回まで議論してきた、脳回路情報として表現される自己に他ならないことに気がつく。ついに、意識と自己が一つにまとまった。
次回はさらに、意識の選択基準について議論を深める。
[ 西川 伸一 ]
意識と無意識
2017年4月3日
最初意識を夢遊病状態と覚醒状態を区別するものとして始めながら、最後はアメフラシにも意識があっていいなどと前回終わったので、混乱させたのではと反省している。
なぜこのような議論になったかを思い出してもらうと、私たちが普通感じている意識のような高次脳機能が関わる現象を、「脳に入力される情報から、それまでの経験をもとに形作ってきた神経系の自己を基準に、一部を選択する」ことと定義したことに起因する。もし意識が入力の選択過程だけなら、最も単純なアメフラシの神経系でも同過程は存在していい。
ただ忘れてはならないのが、選択の基準となる各動物の神経回路に表現されている自己は、単純な回路から複雑な回路まで、多種多様な点だ。神経回路として同じ原理を共有していても、その違いはアメフラシから人間まで途方もなく異なっている。当然、脳神経系が複雑になればなるほど、神経回路に表現された自己自体も複雑になり、それに基づく選択基準も多様かつ複雑になる。従って、脳内に入ってきた情報を自己を基準に選択するという点では同じでも、選択過程は質的、量的に大きく異なる。最初定義した狭い意味での意識はこの選択過程のほんの一部を代表する最も高次な過程に関わっている。
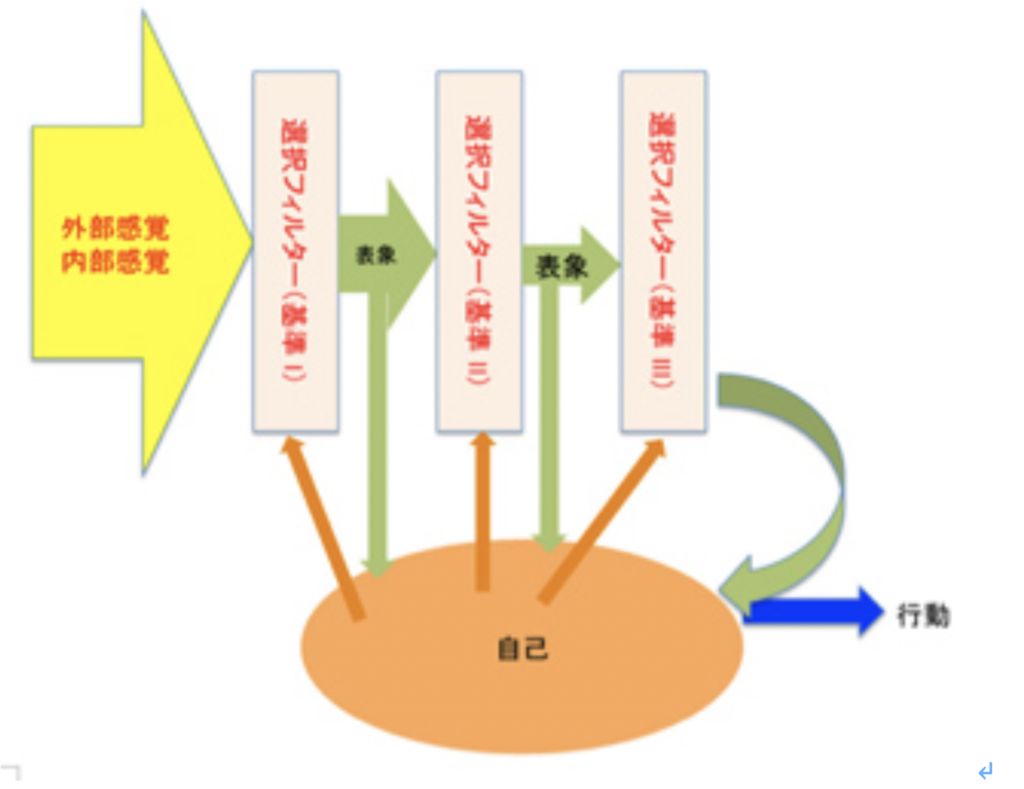
図1:意識を形成する脳内活動。自己の様々な回路に合わせて選択が起こり、選択された表象がまた自己の神経回路に統合される。
図1を使って説明しよう。私たちの脳は多くの情報に曝されているが、基本的には自己の生存に必要な情報だけを選択して他の情報を無視するフィルターを持っている(この傾向がすべての脳回路には備わっており、選択する行為自体を動機付けている)。このフィルタリングを受けた情報が脳内に新しい表象を作る過程が感覚として認識されることが意識だが、神経系の自己全体が反映されたフィルタリングが起こるレベルは多様で、同じ刺激に対しても、様々な表象が選択され、脳の異なる領域に新たな回路が生まれる。こうして選択された様々な表象は、既存のネットワークと統合され、短期的、長期的に脳ネットワークの書き換えが行われるが、最初に定義した意味での意識に上るのはその中のほんの一部になる。すなわち、ほとんどの選択過程は意識されることはない。
もう少し具体的に見てみよう。感覚神経は様々な刺激に反応できる。視細胞は光に、聴覚は有毛細胞への圧力、嗅覚はニオイ物質の嗅覚受容体への結合を神経興奮に転換する。この神経興奮は他の多くの神経興奮とともに統合され表象されるが、この表象は入力とは全く物理的関係を持たず、神経ネットワークの構造変化としてシンボル化されている。まずこの入力に対応する表象を形成する過程でフィルターが存在している。例えばそれぞれの感覚神経細胞レベルで感受できる刺激に限界が存在し、これがフィルターになる。分かりやすく言うと、どんなに頑張っても私たちには紫外線は見えない、また高ヘルツの音波を感じることもない(図2)。
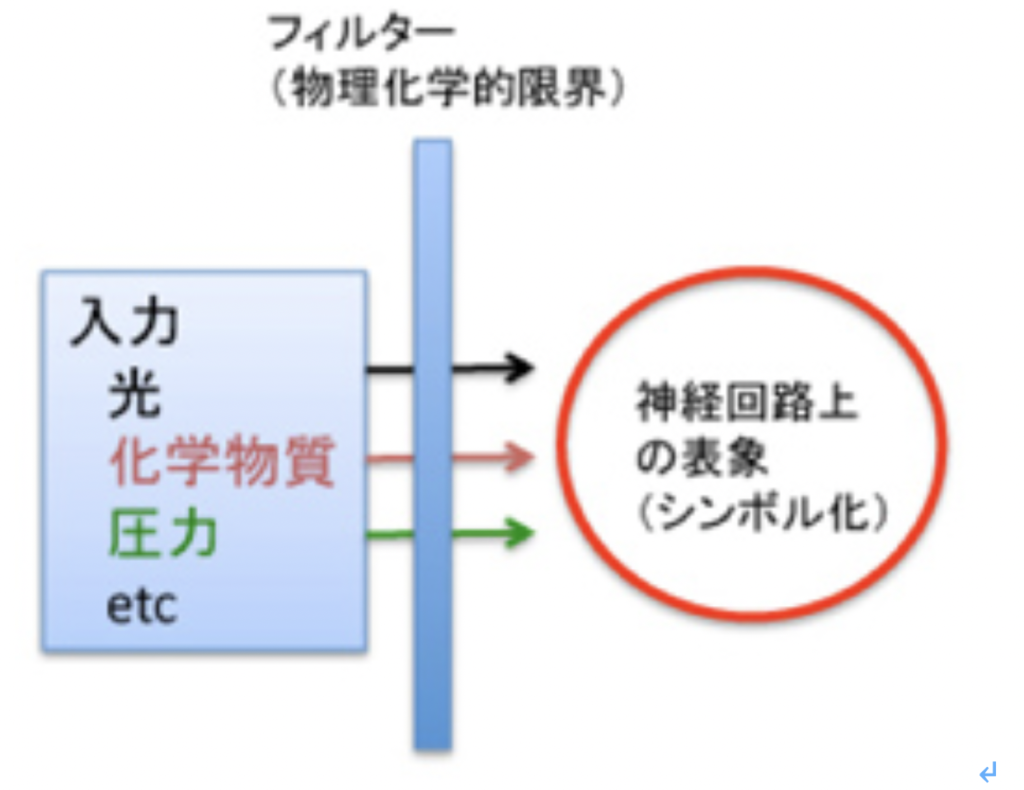
図2:最も原始的なフィルター。それぞれの感覚神経の、刺激に対する閾値は最初のフィルターになる。当然意識されることはない。
次に、アメフラシの水管反射で、エラからの神経回路が、頭からの神経回路の刺激を受けると新たな回路を構成する例からわかるように、他の入力とネットワークが形成されることで、一つの入力だけでは生まれない新しい表象が生まれる。このような表象はしたがって、神経ネットワークの連合の仕方に変化が起こることでもフィルターされる(図3)。
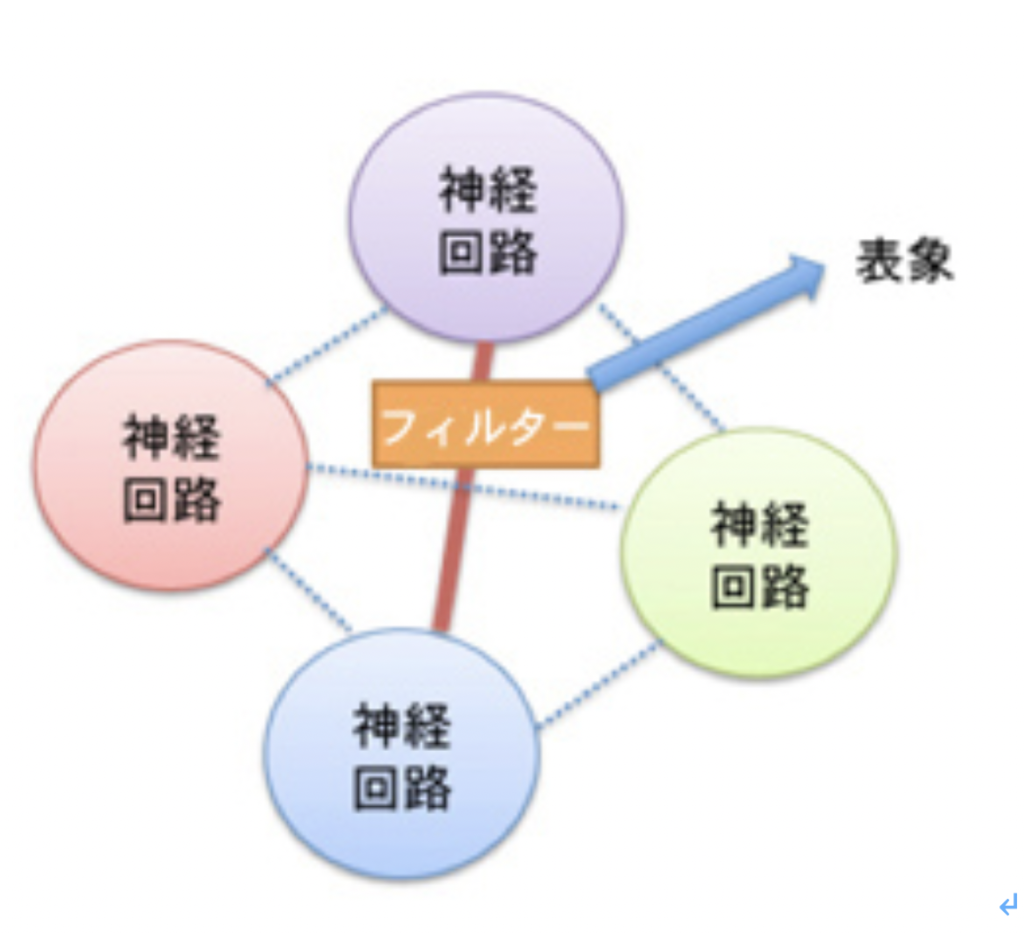
図3:新しい神経回路の連絡路の形成もフィルターになる。
これは単純な回路の例だが、私たちのように複雑な脳になると、連合過程でのフィルターが常に行われていることがもっとはっきり見えてくる。例えば私たちは網膜に入る感覚を、目を動かす動眼神経の運動と統合させることで、空間内の物体を形として背景から区別し、物体の動きを追跡している。すなわち、一つの表象が少なくとも2種類の全く異なる入力を統合することで形成される。すなわちこの統合過程も表象を形成するまでに通過するフィルターと考えることができる。このフィルターの基準となる神経系の自己は遺伝、発生過程で決まるだけでなく、発達途上で経験を通してネットワークが書き換えられることで形成される。
この結果、外来刺激に対する異常な反応は、神経ネットワークの発生異常でも、生後の発達過程の回路形成過程の以上でも起こる。たとえば同じ顔の写真を見たとき、自閉症や統合失調症の患者さんたちが意識に上らせる表象が、普通の人とはずいぶん違っていることはよく知られている。以前私のブログで紹介したことがあるが(http://aasj.jp/news/watch/753)、てんかん発作の原因を確かめるために脳内に埋め込んだ電極を記録に使った研究から、健常児の扁桃体は通常目に強く反応するが、自閉症児は口に反応することがわかっている。すなわち、同じ像を見ても、フィルターされる表象は全く異なる。
このように脳が高次化するに従って、フィルターの基準となる神経の自己も多様化し、フィルターされた表象が書き換える神経ネットワークも多様化している。前回述べたように、人間のような複雑な脳を持つ生物の場合、これらの書き換えは狭い意味で意識される場合もあるし、意識されない(無意識)場合もあるが、行われていることはどちらも様々なレベルの神経的自己を基準にした表象の選択だ。したがって、脳の複雑さが一定レベルに達するまでは、すべての選択は、我々の言う「無意識」下に行われる。一方、特定のレベルを超えると、最初定義した狭い意味の意識過程としての選択が誕生することになる。実際、高度に発達した脳を持つ人間も、生まれてすぐは、入力の選択による表象の形成と新たな脳へ書き換えが起こるとはいえ、おそらく意識はまだ存在しない。例えば両眼競合は起こっても、どちらを選択したかを意識することはないだろう。しかし、成長するにつれ選択結果が意識できるようになる。このことからも、入力の選択による表象が、進化過程、発生過程、成長過程で起こる脳構造の変化に応じて起こり、その一部だけが意識されることがわかる。
ようやく問題が狭い意味での意識に戻ってきた。要するに、意識の基盤にある過程は、ほとんどの動物に存在する神経回路の基本的性質だ。そして、脳の発達があるレベルに達すると、意識が生まれる。ただ意識を可能にする脳機能についてはまだ定説はない。神経回路内での選択過程を認識できる意識に特化した新しい回路が出現したと考える研究者が存在する一方、意識は選択時の基準となる神経回路の複雑化の延長上にあり、意識だけのための特定の回路は存在しないと考える研究者もいる。おそらく現時点ではこの議論を脳科学的にこれ以上進めることは難しいと思う。この結果意識の問題は、もう少し現象論的レベルで議論されることになる。
実際、入力の選択についていえば、無意識下の選択、選択した結果の認識、意識による表象の選択などを、脳科学的メカニズムから離れて現象論的に考えることは現在も普通に行われている。このような様々な意識や無意識、あるいは基準としての様々なレベルの自己、表象選択の動機付けの問題を現象論的に考えた巨人がフロイトだ(図4)。

図4:フロイトのポートレート(出典:wikipedia)
フロイトは精神分析学の父としておそらく最も有名な精神医学者と言っていいだろう。科学者や医学だけでなく、彼は20世紀の哲学に大きな影響を与えた。彼の著作を読んだことがなくとも、彼の用語、「エゴとイド」「リビドー」などの言葉を知っている人は多いのではないだろうか。
彼の生涯はほぼカハールと一致しているが、おそらく彼の精神についての現象論を神経回路に当てはめて考えたことはなかっただろうと思う。しかし普通の人間の行動の傾向、あるいは様々な精神疾患の患者さんの異常な行動を、現象学的ではあっても共通の基盤から科学的に説明しようと努力した。そして、この共通基盤の重要な要素として、「イド、エゴ、スーパーエゴ(エゴイデアル)」、「無意識、前意識、意識」、そして「力動、リビドー」などの概念を提唱した。
これらの概念の詳しい説明は次回に回すが、「イド、エゴ、スーパーエゴ」の概念は、これまで議論した入力の選択基準になる神経回路の様々なレベルの自己に対応すると考えられるし、「無意識、前意識、意識」の概念は、入力選択後の表象が、脳回路を書き換える過程で意識の意味を考えるためのヒントを多く与えてくれる。そして、「力動、リビドー」は、これまで議論してこなかった疑問「意識を維持する力とは何か?意識、無意識を問わず選択、表象、統合に関わる神経回路を動かし、新しい神経系の自己を形成し続ける原動力とは何か?」を考えるための重要な示唆を与えてくれる。
次回は、フロイトが進めた私たちの精神についての現象論について解説して、意識と自己についての説明の最後にしたい。
[ 西川 伸一 ]
フロイトに見る意識と自己
2017年4月17日
意識と自己についての説明を長く続けすぎたので、そろそろ締めくくらなければならない(本当はこの程度の長さで扱える課題ではないのだが)。
普通、意識や自己はかなり高次な神経活動と思いがちだが、これまでの説明で定義次第で様々なレベルの神経活動に当てはまることがわかっていただいたと思う。とはいえ、意識や自己といえば、やはり哲学をはじめ多くの分野で古くから議論されてきた人間の精神活動を思い浮かべるのが普通だ。したがって、締めくくりとして、人間特有の高次の意識や自己について少し考えてみたい。どのように説明すればいいかいろいろ考え、締めくくりにはフロイトを2回にわたって取り上げてみることにした。
今更フロイトを取り上げる意味は本当にあるのか少し迷った。そんな古い話を持ち出さなくとも、現代では、人間の意識や自己について脳科学的に説明した優れた著作を読むことができる。事実、素人の私がここまで脳科学の知識を整理できたのも、アントニオ・ダマシオさんの「デカルトの誤り」「Self comes to mind」「感じる脳」、やクリトフ・コッホさんの「意識の探求」といった本のお陰だと思っている(図1)。

図1:これまで意識や自己を考えるときに参考にしたオススメの著作。最初の3冊はアントニオ・ダマシオ、最後の1冊はクリストフ・コッホ著。
これらの本には多くの脳研究成果が取り入れられており、しかも一般の人にもわかりやすく書かれている。わざわざ解説するより、人間の意識や自己について知りたいと思われる場合、どれか一冊を手にとって読んでもらうのが一番いい。読めば間違いなく満足できると思う。また、上にリストしたすべての著作では実際の脳神経回路と現象を対応させて説明する努力が行われており、症状や行動の現象論的記述と、独断的(に見える)な説明に終始するフロイトを読むよりよほど分かりやすい。
実際、フロイトを読んでも、脳科学の話はまず出てこない。彼の著作にはダーウィンがなんども登場し、進化論の影響を強く受けていることを感じる。もちろん進化の問題だけでなく、種の起源の最終センテンスでダーウィンが全く手つかずの未来の問題として提示した無生物から生物への過程についても興味を持っていたことはまちがいなく、彼の著書の中で「種の起源」に書かれている内容を彼の著作で繰り返しているほどだ(ちくま学芸文庫「快感原則の彼岸」 p162参照、「過去のある時点において、現在もなお想像できない力の影響によって、生命のない物質の中に生命の特徴が芽生えた」)。ところが、カハールも含めて脳科学の話はフロイトの著作にはほとんど登場しない。おそらく当時の脳科学はまず役に立たないと考えていたのだろう。
しかし、神経科学的視点からの説明が全くされていないことは、神経細胞の進化からスタートしてここまで続けてきた私から見ると、本当は都合がいい。脳科学的に考えるにはまだ早い時代であったとはいえ、フロイトは精神現象を共通の基盤に基づいて科学的に説明しようと常に心がけていた。精神現象を、「エス、自我、超自我」、「意識、前意識、無意識」、「リビドー、タナトス」などの共通の要素から分析しており、個別の現象に対する彼の普遍的説明は理解しやすい。このおかげで、フロイトが記述した様々な精神現象と、それについての彼の説明を、もう一度神経科学の視点から整理し直すことができる。一方、これまで私自身は人間特有の高次な意識や自己について議論を避け、神経ネットワークが複雑化する延長に人間特有の精神活動もあるとして済ませてきた。この認識は間違っていないと思うが、複雑化することで何が可能になったのかは全く手つかずのままだ
一方、フロイトの興味はもっぱら人間の意識だ。従って、自己や意識についての彼の考えを現代の脳科学を知った上で見直すことで、これまでなおざりにしてきた人間特有のレベルの意識や自己の発生に関わる過程についてもヒントが得られることも期待できる。これが、フロイトを選んだ理由だ。
フロイトの考えを紹介するための著作として、今回はフロイトの短い著作「自我とエス」(竹田青嗣編、中山元訳、自我論集、ちくま学芸文庫)を選んだ。これは後期の著作で、一般人に向けた読み物というより、総説論文に近く、短いとはいえ彼の考えが詰まっており、また現代脳科学からも扱いやすい。
例えば意識や自己の発達過程について、フロイトは次のように述べている。
「個人の発展の最初期の原始的な口唇段階においては、対象備給と同一化は互いに区別されていなかったに相違ない。のちの段階で性愛的な傾向を欲求として感じるエスから、対象備給が生まれるようになったと想定される。最初はまだ弱々しかった自我は、対象備給についての知識を獲得し、これに黙従するか、抑圧プロセスによってこれから防衛しようとする。」
「少年の成長について簡略化して記述すると、次のようになる。ごく早い時期に、母に対する対象備給が発展する。これは最初は母の乳房に関わるものであり、委託型対象選択の原型となる。一方で少年は同一化によって父に向かう。この二つの関係はしばらくは並存しているが、母への性的な欲望が強まり、父がこの欲望の障害であることが知覚されると、エディプス・コンプレックスが生まれる。」
この2つの文章には、意識や自己という言葉は使われていないが、フロイトが考えた意識や自己の形成過程に必要な要素が凝縮している。また私がこれまで人間に限定せず見てきた自己や意識について述べてきた重要な要素も含まれている。従って、目標はこの文章を理解することだ。ただフロイトを読んだことがない読者もいるだろうし、彼の文章には一般には使われることのない単語も多く使われているので、まずフロイトの意識や自己に関する考えについて総論的に要約する。とは言っても、ここで述べることは全て私の独断的なフロイト理解であることは断っておく(フロイト解釈で議論する気は毛頭ない)。
まず意識、無意識から始めよう。フロイトというと、「無意識」がすぐに思い出される。しかし彼の著作では無意識については詳しく述べているものの、「意識する」とは何かについて、ほとんど明確にしていないように思う。この論文の中でも「<意識されているとは、まず純粋に記述劇な用語であり、最も直接的で確実な知覚に依拠するものである」と、意識されているものについて簡単に済ませてしまっているが、肝心の「意識とは何か?」についての彼の考えは明確ではない。おそらく彼の理解も、私たちが一般的に意識という言葉で使っているのと同じ内容、あるいは夢遊病を例に定義した意味とあまり変わっていないように思う。
これはフロイトの問題ではなく、意識の厳密な定義が現代でも難しいためで、フロイトの時代にはまだ「両眼競合」のような実験結果も知られていなかった。しかし彼の著作を読むと、彼も私たちと同じように意識を様々な知覚から生まれる脳内の表象を認識して選び取る過程として捉えていたように思う。
私たちはともすれば「意識 vs無意識」と対立的に提示してしまうが、そもそも両者を比べること自体難しい。よく考えてみると意識とは多くの表象の中から特定の表象を知覚する「過程」を意味することが多い。一方、無意識の場合、「無意識する」のようには決して使わず、常に意識過程の外にあるという「状態」を指すことが多い。従って、「意識されるものvs無意識のもの」と対置するのは不自然で、「意識されるものvs意識されないもの」の方が自然だ。すなわち、意識はより過程に関わる言葉として使われることが多く、もう一方は状態を指す言葉として使われる。
フロイトの興味はもちろん人間の心の状態なので、意識されるものについてはあまり重視していない。代わりにフロイトは心の状態の最も大きな部分をしめる、無意識に向かうことになる。
誤解を恐れずフロイトの無意識を「エイヤ!」と意識との関わりから分類してしまうと、無意識とは、
の3種類に分けられる。
最初の2つについては、フロイトは「無意識は潜在的に意識できる状態と一致するもの」と述べて、起きている時にいつでも意識できたのだが,たまたま意識過程で選ばれなかったり、忘れてしまったものとして簡単に済ましている。これを脳科学的に見れば、感覚を通して入ってくる膨大な入力の中からほんの一部の表象を選んで神経ネットワークの書き換えていることに相当する。
ところが、3番目の「意識させてもらえない」無意識は、フロイトの言う「抑圧された」無意識で、私もこれまで全く議論してこなかった。この抑圧された無意識が何を意味するかについては、最初に引用した文章によく書かれている。文章に出てくる備給という言葉はおそらく馴染みがないと思うが、Cathexisの日本語訳で、心的な関心を向けることを意味する。私自身はこの大げさな単語を見た時は、意識する表象を選択する過程に、具体的な方向性と必要なエネルギーを与えることと読み直している。
この文章でフロイトは、父親についての表象が生まれることで、幼児時代に本能的に母親の乳房に向いていた意識過程、すなわち母への性的欲望が抑圧され、無意識(すなわち意識過程から除外された状態)へと追いやられると述べている。すなわち、成長に伴い、本能とは異なる判断基準が自己の中に芽生え、これが母親への性的意識を無意識へと抑圧する。この抑圧された結果としての無意識は、毎日の感覚の中から意識下、あるいは意識外へと選択される無意識とは全く異なり、抑圧を取り除かない限り無意識から意識へと上らせることは難しい。すなわち、抑圧される無意識は、意識過程で選択が行われるという意味では同じだが、両眼競合で見られるような選択とは異なり、高次のレベルで抑制され続けている心的状態と言える。
このように、最初の2つの無意識と、3番目の無意識は質的に異なっていることから、フロイトは前者を前意識、後者を無意識と名付けている。このように、無意識に様々な種類があるということは、意識と無意識を分ける時の基準になる自己にも様々なレベルが存在することになる。この基準としての多様な自己を、フロイトは「エス、自我、そして超自我」という言葉で対応させた。
図2:フロイトの「自我とエス」より転載。
図2は「自我とエス」に掲載されている、自己の精神についてフロイトが描いた図式をほぼそのまま転載したものだが、この図を見ながら彼が自己をどのように考えていたのかもうすこし見ていこう。
線で囲まれた部分が個人の精神的自己全体を表しており、この中に前意識、抑圧された無意識という2つの状態も含まれている。重要なのは、自己の中の表象を選び認識する過程である意識は、自己の枠外に置かれていることで、これにより意識が自己の中に存在するコンテンツではなく、より過程に近い物であることがうまく表現されている。
では、エスや自我とは何か?引用した文章の中でフロイトが、「最初弱々しかった自我」と表現しているように、最初から自我は個人の心に存在するのではなく、生まれてから経験を繰り返すうちに心の中に芽生えてくる物として考えられている。すなわち、経験を通してのみ形成される新しい選択基準と言っていいだろう。
一方、エスは生まれた時から存在し、母への本能的な性的愛情も最初から存在するエスの表れと考えられている。すなわち、フロイトのエスは人間が生まれた時に持っている脳神経ネットワークが生きるために最初から備えている様々な本能と言っても良いだろう。
私自身はこれまで自己についてこれほど詳しく区別せず、脳神経ネットワークが複雑化した結果、選択基準となる自己も多様な表象を持っており、一人の個人に様々なレベルの自己が存在するとして済ませてきた。しかし、生まれてから物心着くまでの一定期間、人間には狭い意味での意識とその記憶は存在していない。従って、記憶や自己の意識は、生後の成長期に形成されると直感できる。この直感は、「自我からその核としての知覚システム(意識)が形成される(図1に表現されている)」というフロイトの考えと同じだ。
誕生時に持ち合わせた脳神経ネットワークの活性を「エス」、この中で成長に応じて高次化する意識過程で選ばれた表象取り込んで脳神経ネットワーク(エス)の中に新たに形成した回路を「自我」考えることはできないだろうか。脳科学的に、純粋に生後の経験だけで生まれる「自我」を完全に分離できるかわからない。フロイト自身も、「エスは、知覚—意識システムの媒介のもとに、外界の直接的な影響を受けて変化する。自我はこのエスの一部である。・・・・」と、本当は両者を完全に分離することはできないと考えている。ただ、経験を通して自伝ともいうべき基準を人間は作ることができるおかげで、新しいレベルの意識が人間には可能になっている。この感情や本能(エス)の上に、自伝を書くメカニズムの解明が、人間とは何かを理解し、次の情報媒体「言語」を考えるための第一歩になる。
期待通りフロイトをたどることで、これまで手がつかなかった人間特異的なレベルの自己や意識の問題について考えることができてきた。まだ、母親へ向かう性的本能を抑圧するのは自我か?フロイトの言うもう一つの自我、「超自我」とは何か?そして意識という過程にエネルギーを与える力は何か?など、フロイトの考えの解説が残っているが、少し長くなりすぎたので、今回はここで止める。次回、最後はフロイトの描いた図1を、現代的な新しい視点で書き直すことで、最初に引用した文章を完全に理解して終わりたいと思っている。
[ 西川 伸一 ]
フロイトの意識と自己
2017年5月1日
前回・今回とフロイトの考えを紹介しているが、彼の文章を題材に、高次の意識や自己を説明することが目的だ。そこで、前回引用したフロイトの文章をもう一度掲載しておく。
「個人の発展の最初期の原始的な口唇段階においては、対象備給と同一化は互いに区別されていなかったに相違ない。のちの段階で性愛的な傾向を欲求として感じるエスから、対象備給が生まれるようになったと想定される。最初はまだ弱々しかった自我は、対象備給についての知識を獲得し、これに黙従するか、抑圧プロセスによってこれから防衛しようとする。」
「少年の成長について簡略化して記述すると、次のようになる。ごく早い時期に、母に対する対象備給が発展する。これは最初は母の乳房に関わるものであり、委託型対象選択の原型となる。一方で少年は同一化によって父に向かう。この二つの関係はしばらくは並存しているが、母への性的な欲望が強まり、父がこの欲望の障害であることが知覚されると、エディプス・コンプレックスが生まれる。」
前回、フロイトの意識、前意識、無意識の区別、あるいはエス、自我、対象備給などについて説明したので、この文章もだいぶわかりやすくなったのではないだろうか。
今回は私自身がこの文章をどう読んでいるかもう少し突っ込んで紹介しながら、最後に残っている超自我や、私たちの意識や自己を動かしている力動について説明したいと思う。
口唇段階の本能で動く脳がエスの始まり
生まれたばかりの私たちは「原始的な口唇段階」、すなわち食べ物を得るため母の乳房を探るという本能により動かされる段階にある。実際、この段階の神経回路の特徴を小児科では原始反射と呼んでおり、口唇に関わる反射が多い。例えば口に物が入ると吸い付く反射、口唇に触れるものを追いかける反射などが有名だ。フロイトが「口唇段階」と名付けたのもこのような古くから知られる赤ちゃんの行動に基づいている。フロイト流にいえば、この段階の精神はほぼエスに等しいと言える。
従ってエスをもう少し実体的に表現すると(対応させること自体は意味がないが、理解のためにこのような対比を今後も行う)、外界からの刺激を経験しない、発生が終わったばかりの脳内神経ネットワークと言える。この脳には自分の生命を守り、外界からのストレスを避けるためのメカニズムが進化の過程でプログラムされている。
幼児の成長に伴う脳の書き換え(同一化)
生まれるとすぐ(実際には生まれる前から)、動物は様々な感覚システムを通して外界から刺激を受け、その一部は高次の意識とは無関係に、できたばかりの脳ネットワークに介入し、ネットワークを書き換えることができる。以前議論したようにこの書き換えは、膨大な入力の中から私たちの脳がその時点で形成している「自己」の基準に従って特定の入力を選択し、神経ネットワークに記憶する(書き換える)ことで進んでいく。これが広い意味での意識だが、この書き換え課程は、フロイトの文章の中の「同一化」とほぼ同じと考えていい。生まれたばかりの「口唇段階」の自己(この場合エス)では、行動のほとんどが母の乳房へと向く様に脳はプログラムされている(対象備給)。しかしこの単純な行動を通して、様々な外界からの感覚を経験する。最初は口唇感覚に限られていたものも、皮膚感覚や匂い、音、そして最後に視覚というように入力装置は複雑化するとともに、入力量は急速に拡大し、新しい経験で私たちの脳のプログラムは徐々に書き換えられていく。この初期段階での入力を選択する自己の基準はエスであり、生まれた時点で備わっている本能的自己だ。もちろん、入力が選択され神経ネットワークの自己が書き換えられるとき、エス=本能的自己も書き換わっていく。このことは、先に述べた新生児の原始反射が発達に伴い消失することからわかる。代わりに、より複雑な外界からの刺激を選択し、本能的自己を書き換える過程で、ハイハイなどの新しい機能が発達する。
自我と高次の意識の誕生
問題は、このような神経ネットワークの書き換えの延長に、高次の意識や自我が発生するかで、この点が意識とは何かをめぐる議論の一番重要な論点だ。おそらくフロイトは自我がエスの単純な延長にあるのか、それともエスとは別の自我を受ける構造が発展するのかあまり気にしていなかったのではないだろうか。前掲の文章で彼はエスから自我が芽生えると述べており連続的に考えていた様に見える。ただ「自我とエスの区別は、原始人だけではなく、はるかに単純な多数の生物にも認めることができる。この区別は、外界の影響の必然的な表現だからである」と、最初の脳ネットワークの自己であるエスと、外界の感覚入力を通して形成され成長するもう一つの自己である自我を分けることの重要性も認識していた。すなわち、エスの中に、エスとは区別された新しい自己、自我を発生させる入力処理機構が必要になる。
しかしこの区別は決して前回、図1で示したような固定した区別ではない。抑圧された無意識もエスの一部として記憶されると彼が考えた様に、エスはキャパシティーの大きな、自我も無意識も全て包含することができる、変化し続けるフレキシブルな構造だ(本来脳とはそのようなものだ)。このような構造の中にエスとは別の自我が形成されるのは、生後間も無く経験する感覚器を通した様々な入力が、高次の意識や自我を持つ動物の進化で生まれた、それまでとは別のまったく新しいチャンネルを通して処理され、表象されるからと考えられる。
自我を形成する新しい入力チャンネル
実際、口唇期の感覚の中心となる触覚と比べると、最後に発達する視覚認識は、もともと多くのチャンネルを必要としている。私たちは目に入ってきた人間の姿を認識するとき、網膜の視細胞からの入力を、色や単純な形、あるいは物体の動きといった要素として別々に表象する。この過程はほとんど意識されることはない。これらの要素がイメージとして知覚されるためには、この要素を今度は過去の経験も合わせて上から(トップダウン)統合する必要があり、このトップダウン過程は意識される。このように、私たちの脳は本来多数のチャンネルを使って、感覚入力を処理するようにできている。そして、生後経験し続ける感覚入力を統合する際の特別な領野が用意できれば、そこに自我、あるいは新しい自己を形成することが可能になる。これを意識をコードする領域と言っていいのかもしれない(あるかどうかは議論が多いが)。
ではこの新しい領野にどの入力が選択され、記憶され、自我を形成するのだろうか。最初存在する自己の基準はエス、すなわち本能的自己により提供される。生きる本能が乳房を追いかける反応として現れることを思い出してほしい。しかし、エスのレベルで選択されたどの表象が、生まれたときにはまだ白紙状態の自我の書き換えに利用できるのかよくわからない。口唇の触覚を通して経験される感覚はおそらくそれほど複雑でなく、ほとんどの入力が最初の自我の形成に使われるのかもしれない。こうして一度でも自我の形でエスとは別の自己が成長し始めると、この自己を基準に自我回路は書き換えられ続け、自我レベル、すなわち意識される自己が成長する。ただ、入力は意識され、自我の書き換えだけに用いられるわけではない。エスも様々な入力により書き換えられるため、生まれたばかりの状態を代表する様々な原始反射は消失する。
エス・自我・意識のまとめ
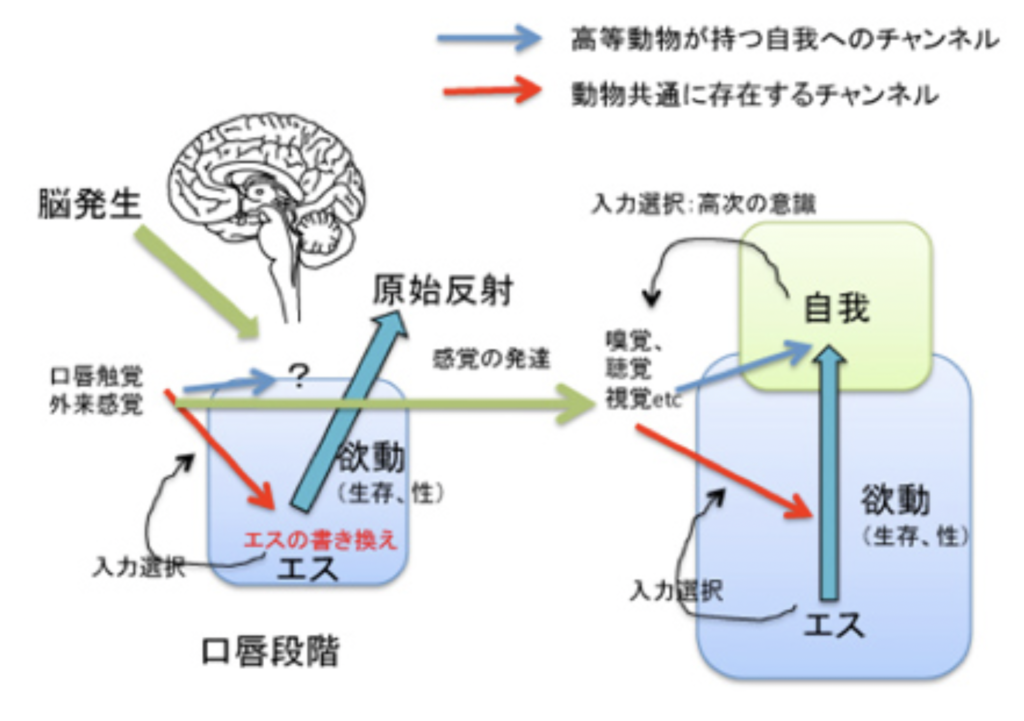
図1:乳児の発達期の感覚入力と自己の書き換え
これまで議論したことを図にしてみた(図1)。口唇期では外界からの入力はエスとしての自己を基準に選択され、エスの書き換えが行われるとともに、萌芽的自我の形成とその後の書き換えに使われる。こうして自我という新しい自己の基準が形成されると、その後は新しい自己の基準として、高次の意識過程を可能にする。この結果、私たちは「物心がつく」、すなわち感覚器から入ってくる経験を意識し自己の形成に使う。
フロイトは、知覚から生まれる前意識が意識されると、こうして得られた表象は全て自我の形成へと向かうと述べているが、実際私たちの自我が意識的に知覚される入力を選択する唯一の基準になると考えれば当然のことだ。自我が高次の意識の基準であり、それを基準に選択された表象が自我を書き換える。以上が、最初に引用したフロイトの文章の現代風読み替えになる。おそらくみなさんも、引用した文章でフロイトが伝えたかったことは、概ねわかってもらえたのではないだろうか。
しかしまだ、自我により抑制された無意識や、超自我についての説明が残っている。これらの概念は、私たちを自我やエスに基づいて行動へと駆り立てる精神的エネルギーと関わっており、私自身も全く議論してこなかった。
快感を得るための欲動
フロイトは、このエネルギーの源が、「快感」が満たされるまで止むことのない欲動であるとはっきりと述べた。この欲動の起源についてフロイトは、最も重要なのは生存への欲動と、繁殖への欲動で、両者に共通する最も大きなエネルギーの源が性的欲動、リビドーであるとした。このリビドーという用語は、その後ユングにより欲動一般を指すものとして定義し直される。しかし、当時自己とは何かなどと議論していた宗教家や哲学者を尻目に、私たちの脳は快感を追求するようプログラムされているおかげで行動するエネルギーを得ていると、ズバリ指摘したのは、フロイトの最も偉大な業績の一つだろう。
余談になるが、「快感原則の彼岸」とタイトルのついた小論文で、彼はこの欲動の生物学的起源について進化論に即して議論している。単細胞動物の生殖や性、あるいは多細胞動物の始まりと「個体の死」の始まりといった当時の生物学の知識を総動員して、「生きることへの欲動」「生殖のための欲動」そして無限に続く生殖細胞系列を守るための「死への欲動」などが、精神を動かすエネルギーのルーツであると述べている。言葉を変えると、私たちの精神行動エネルギーが、生命として必然的な帰結であることを述べている。
欲動の中で最もわかりやすいのは「生存への欲動」だろう。例えば新生児期を考えると、口唇段階でのこの欲動は空腹を満たすために必須のメカニズムだ。欲望が満たされるまで、子供は母の乳房を求め続ける。男の子になると、この生存への欲求に、男性が本能的に持っている女性への性的欲動が混じる。いずれにせよ、感覚器が発達し、複雑な感覚入力により自我が複雑化するにつれ、欲動の表現も複雑になる。しかし、そのエネルギーのルーツを辿れば、必ずエスの中に組み込まれた生存欲、性欲を生み出す脳構造に帰結する。これを
「個人の発展の最初期の原始的な口唇段階においては、対象備給と同一化は互いに区別されていなかったに相違ない。のちの段階で性愛的な傾向を欲求として感じるエスから、対象備給が生まれるようになったと想定される。」
とフロイトは表現している。
エディプス・コンプレックスの起源
この欲動のエネルギーおかげで幼児は行動し、外界と持続的なコンタクトを維持することができる。最初は原始反射を繰り返しながら、生後徐々に発達する五感を通して経験される母親の表象は常にアップデートされ、幼児期の自我の書き換え(すなわち同一化)の基準を提供する。もちろん人間社会では通常子育てには父親など母親以外の人間の参加があるが、母親以外が表象され自我の書き換えに参加するのは、五感、特に視覚が完成し、母親以外の人間が意識されるようになってからのことだろう。したがって、最初の自我は母親の表象を中心に形成されており、それ以外の表象は常に母親との対比で自我の書き換えに参加する。
これは私の想像で当時のことを覚えているわけではないが、子供を可愛がるのは母親だけではない。父親も子供をあやそうと抱きかかえるとき、子供は母親から一時的にでも引き離されると感じるかもしれない。もちろん父親には母親に期待する乳房はない。当然母とは異なる父親の表象が生まれ、自我を書き換える過程で、母親から引き離す新しい競争相手として父親をイメージしても不思議はない。
フロイトは、このとき形成される母親をめぐる競争相手としての父親と自我との関係を、精神の発達に最も重要な要因であると強調し、
「ごく早い時期に、母に対する対象備給が発展する。これは最初は母の乳房に関わるものであり、委託型対象選択の原型となる。一方で少年は同一化によって父に向かう。この二つの関係はしばらくは並存しているが、母への性的な欲望が強まり、父がこの欲望の障害であることが知覚されると、エディプス・コンプレックスが生まれる。」
と述べている。
「不快」を行動のエネルギーにする超自我
しかしもし子どもの行動を支配するエネルギーが「快感原則」であるとすると、「不快」がなぜ精神を動かすエネルギーになるのかが問題になる。
もちろん動物は「不快」な経験を記憶し、それを回避する脳回路を開発している。当然この書き換えは、自我の書き換えを伴う。だとすると、「不快」の原因である父親を、避けることで母親との「快」の関係を維持する以外に「快感原則」を守る方策はない。おそらく、多くの動物で子育てにオスが関与しないのは、子供への愛情が発達しないのではなく、子供にとって父親が「不快」でしかないことが原因かもしれない。
一方人間を見ると、乳児が父親の「不快」な介入を避けることは難しい。従って、不快を我慢する能力を身につけるか、あるいは積極的なエネルギーへと変換できる自我を開発する必要がある。この「快感原則」と取引できる自我の形成こそが、人間の利他的行動や、道徳の起源だと思う。この「不快」を選ぶという人間の特殊性を、フロイトは無意識と、超自我という概念で説明しようとした。
この問題提起は
「最初はまだ弱々しかった自我は、対象備給についての知識を獲得し、これに黙従するか、抑圧プロセスによってこれから防衛しようとする。」
という文章に見ることができる。
すなわち母親や父親のイメージが明確になってくると、それまで「快感原理」のみに動かされていた行動に矛盾が生じ「不快」を生むようになるが、残念ながら両親に守られている身分では、「不快」を避けるという解決自体が不可能であることが理解される。これが、「父がこの欲望の障害であることが知覚されると、エディプス・コンプレックスが生まれる。」と彼が語る、エディプス・コンプレックスだ。しかし、このコンプレックスは、行動で解決できる代物ではない。このため、不快を避けるのではなく、自分で解決する必要があり、そのために自我を大きく書き換え、自我の領域にこの問題を解決するもう一つの領域、「超自我」が発生する。
この超自我の誕生により、「自我理想(超自我)はそもそもエディプス・コンプレックスの抑圧というこの急激な転換によって成立する」と彼が語るように、「不快」を「快感」へと転換させるエネルギーが生まれる。
すなわち、「快感原理」自体が私たちを動かす本来のエネルギーであっても、この原理を維持するため、不快な経験を快感原理と妥協させる能力、すなわち超自我を人間は獲得した。したがって、フロイトはあらゆる道徳的、宗教的動機は全てこの超自我の仕業であると喝破して見せた。
このように、エス、自我、超自我、前意識、意識、無意識、欲動、リビドーというフロイトの用語は、現代の脳科学から見ても十分説得力があり、さらに人間特有の高次な脳機能を考える時、色褪せることのない道標になってくれる。この中で、超自我として表現される機能は人間特有の機能だろう。従って、彼の超自我の機能の解明は、21世紀脳科学の最大の問題の一つと言っていいだろう。
これでフロイトは終わるが、次回は整理の意味で、私たちの精神のエネルギー、情動について現代の脳科学がどのように考えているのか、幾つかの総説を基礎に述べてみたい。
[ 西川 伸一 ]
快感原理
2017年5月15日
「ケセラセラ、なるようになる」と楽しく生きている人もいると思うが、多かれ少なかれ、だれでも自分の行動原理を持っている。例えば宗教や道徳は多くの人にとって行動を決めるときの強い原理として働いている。実際「あなたは行動に当たって何を最も大事に思いますか?」とあらためて聞かれて、「快楽を求めて生きています」と、ドンファンを気どる人はほとんどいないはずだ。私も、「余生は若い人のために」とか「患者さんの助けになれば」とか答えるのが普通だが、答えた途端なんとなく偽善的だなと後ろめたい気持ちに襲われる。
人間には従うべき行動原理があるというドグマに対し、私たちの行動のエネルギーの最も重要な要素は快感原理に他ならないと断定したのがフロイトだ。私たちの快楽を追求するために行動し、嫌なことを避けるために行動しているだけだと建前を切り捨てた。
しかしこのことに気づいたのはフロイトが最初ではない。同じ趣旨のことを17世紀オランダで活躍した偉大なユダヤ人スピノザは彼の著書「エチカ」の中で、人にとっての善悪は結局好きか嫌いかの感情でしかないと述べている。
図1:スピノザの肖像
スピノザはポルトガルからオランダへ移住してきたユダヤ人、マラーノの家族に生まれた。デカルト、ライプニッツと並ぶ17世紀の近代を開いた哲学者。「エチカ」は、科学や人間を考えるとき、「神学・政治論」は自由を考えるとき、今も新鮮だ。(出典:wikipedia)
その幾つか引用すると(全て岩波文庫、畠中尚志訳)、
「我々をしてあることをなさしめる目的なるものを私は衝動と解する。」
「全及び悪の認識は我々に意識された限りにおける喜びあるいは悲しみの感情にほかならない」
「我々は速やかに出現するだろうと表象する未来のものに対しては、その出現の時が現在からより遠く隔たっていると表象する場合よりもより強く刺激される」
「善及び悪の真の認識が感情である限り、それから必然的に欲望が生ずる」
「喜びから生ずる欲望はその他の事情が等しければ、悲しみから生ずる欲望よりも強力である」
などなど、要するに「善は快楽、悪は不快」とまで言ってしまうと、17世紀にかなり自由な国だったオランダとはいえ、キリスト教支配のヨーロッパでは禁断の思想となったのは当然だ。
前置きが長くなったが、これまで長々と述べてきた「意識・自己」についての話の最後は、快楽がどう私たちの行動を支配しているのかについての最近の研究を紹介して終わる。スピノザやフロイトも脳の快楽中枢について研究が行われる日が来るとは想像だにしなかっただろうが、この分野の第一人者Berridge とKringelbachの共著で書かれた2編の総説 (Kringelbach & Berridge, The pleasure circuit found in the brain. Scientific American , p40, 8月号、2012、 Berridge & Kringelbach, Pleasure system in the brain, Neuron, 86, 646, 2015)を読むと、面白い研究が進んでおり、これを読めばフロイトも大いに興奮しただろうと思うので、これら総説の内容を私なりに脚色しながら紹介したい。
電極を挿入して局所的に脳を刺激する治療法(脳深部刺激療法)は現在パーキンソン病を中心に様々な疾患に利用されている。しかし脳の病気を電気刺激で治療する可能性追求の歴史は古く、1938年イタリアで統合失調症に対して行われたいわゆる電気ショック療法が最初だ。私が医学生の頃、精神科ではこの治療法は広く行われていた。効果はあると習ったが私自身は現場に立ち会ったことはない。その後、様々な向精神薬の開発が進み、通電時の身体的・精神的苦痛を懸念して、この治療は下火になった。
代わりに外科的に電極を脳に埋め込み、留置した電極を介して微小電流を脳内で流す脳深部刺激治療が開発され、現在パーキンソン病の治療などに広く行われるようになっている。この方法の開発研究の初期に、重いうつ病治療のために視床下部や側坐核周辺の脳幹部に電極を挿入された患者さんが電気刺激を受けている間だけだけうつ症状が改善し、笑うようになることが報告された。驚くことに、この治療を受けた一人は、刺激スイッチを自分で押すことを許可されると、なんと3時間に1500回もボタンを押し、一種の中毒症状を示したことが報告される。この結果は、脳の中に快楽中枢があることを示す証拠として大きな話題になった。
一方実験動物ラットの中脳辺縁ドーパミンシステムを刺激する実験からも、快楽中枢が存在することが示唆され、この実験は快楽電極(Pleasure electrode)実験と呼ばれた。この部位の刺激はラットに快感を与え、ラットを条件付けるための褒美として用いることができることがわかった。例えばラットを大きな飼育箱に入れ、4つのコーナーのうちどこに集まるかを、電気刺激で快感を与えることでコントロールすることができる。さらに驚くのは、ラットが自分でレバーを押して電気刺激をオンにできるようにすると、ラットは中毒に陥ったように自発的にレバーを押し続けるようになることが示された。この刺激中毒に陥ったラットは、食べるのも忘れてレバーを押し続け、なんと1時間に1000回もレバーを押し続けたことが記録されている。
図2:最初に考えられた快楽中枢システムの伝達経路(赤線で示している。)。被蓋からの経路はドーパミンを最も重要な神経伝達分子として使っている。
この結果は他の研究グループによっても追試が行われ、図2の赤線で示した回路が脳の快楽システムを伝達し個体の行動を調節する回路として考えられるようになった。この回路の起点の被蓋にはドーパミン作動性の神経が存在しており、ドーパミンこそが様々な領域の脳細胞に褒美として使われ、行動を支配するニューロトランスミッターであるする「ドーパミン快楽説」が通説になる。実際、中毒になるほどの行動支配力が電極刺激で得られるなら、このシステムが快楽中枢として行動を支配するとするこのドグマは説得力があった。
しかしドグマは常に破られる。このドグマを疑って、快楽による行動支配のメカニズムについて新しいモデルを示したのが、BerridgeとKringelbachのグループだ。研究の歴史はすべて割愛し、快楽による行動支配が現在どう理解されているのかを、彼らの説に従って見ていこう。
彼らは人間や動物の注意深い観察を基礎に、快楽反応が実際には、3種類の過程、Wanting(モチベーション:欲)、Liking(好み、満足)、それにLearning(学習)から構成され、それぞれの過程は別々の神経メカニズムで調節されていることを明らかにした。Learningは一般的な学習のことで、快楽の基準を形成するためには必須だ。例えば、子供の頃には食べられなかった食品も、学習によりうまいと感じる。このように学習は快楽行動の形成に必須だが、今回は説明を省き、WantingとLikingについてのみ説明する。
BerridgeとKringelbachが快楽を感じている指標として最も重視しているのがlikingと彼らが呼ぶ行動で、人間の赤ちゃんからネズミに至るまで、甘いものを与えると、顔の緊張が取れて、口を少し開けて締まりのない表情になるのを観察することができるが、これに相当する。すなわち、満足感が表に出てしまうおかげで、liking反応が起こっているかどうか判断できる。(Neuronに発表された総説はウェッブからフリーアクセスなので是非写真を見て欲しいhttp://www.sciencedirect.com/science/article/pii/S0896627315001336)。
快楽行動が、WantingとLikingと2種類の神経科学的にも別の過程から構成されていることを確認するため、「快楽反応の主要因」と考えられていたドーパミンの分泌を低下させた動物に砂糖水を与える実験が行われた。
Wantingを砂糖水の消費量、Likingは顔の表情を指標として調べると、ドーパミン産生を低下させたマウスでは砂糖水の消費量は極端に低下するが、甘い砂糖水を飲んだ時はlickingを示すことがわかった。一方ドーパミンの脳内濃度が高いマウスでは、砂糖水を飲もうとする行動が促進されるが、飲んだ後のlikingの表情は示さない。この結果から、これまで快楽回路として考えられてきたドーパミン作動性の回路はWanting、すなわち行動への欲望を高めるのに関わっており、甘い水を快楽と感じることには関わらないことを示している。
このことは、先に述べた深部刺激のレバーを押し続けるうつ病の患者さんでも確認された。すなわち、彼はボタンを押し続けていても、一言も「いい気持ち」とは語っていない。ただ、追い立てられるように行動を起こしているだけだ。おそらくこの快楽が達成できていないことが、ボタンを押すのを止められない理由かもしれない。このように、欲動と、それによる快感は分離することができる。
さて快楽行動に関わる2つの過程を分離して調べるための指標が決まると、次にlickingに関わる脳領域を特定するため様々な場所に電極を挿入し、その細胞を刺激(この研究ではエンケファリン(麻薬物質)が注射されている)することでlikingの表情が現れるかどうか調べることができる。脳内の様々な場所に電極を挿入してliking反応が誘導できる部位が特定され、これらの領域をhedonic spot(快楽スポット)と名付けた。
このhedonic spotという名前からわかるように、例えば快楽を支配する中枢に相当する大きな領域が存在するというものではなく、様々な領域の中にラットの場合1mm立方ぐらいの大きさの小さな独立したスポットが集まる領域が存在することが分かった。また、hedonic hotspotが集まる領域は決して一箇所ではなく、傍小脳脚核、腹側被殻、側坐核、眼窩前頭皮質、島皮質がhedonic spotが集まっている代表的な領域だ(図3で黄色のスポットとして示している)。それぞれの領域は相互に関係しているが、例えば食事の満足で見ると、側坐核内のhedonic spotsと腹側被殻内のhedonic spotsが最も大きな役割を果たしていることが明らかにされている(図3:黄色の矢印)。それぞれは、眼窩前頭前皮質にあるhedonic spotとネットワークを作っている。これらの領域を障害する研究から、3つの領域の中では腹側被殻の障害が最も強い影響が見られることが分かっている。すなわち、hedonic spotsはネットワークで結ばれ、それぞれが異なる機能を持っていることが予想される。
図3:側坐核前方と腹側被殻を中心とするhedonic ネットワーク。説明は文中。実際の実験はラットで行われ、図3はそれを人の脳にマッピングし直している。快楽を与えてどこが興奮するか機能的MRIを用いて調べ、人間でのhedonic hotspotのマッピングも行われており、ラットとの結果と重なる。
さて、図2に示したように、Wnating(欲動の回路)を支配している最も重要な分子はドーパミンだったが、hedonic spotでは麻薬に反応する受容体が重要な働きをしている。ただ、麻薬に対する反応が常に快楽をもたらすわけではない。例えば、最も研究されている側坐核では、麻薬や大麻などの刺激に対する受容体と体内麻薬分子がホットスポットの興奮に関わり、liking反応を誘導する。しかし同じ側座核でもhot spotを外れると、同じ刺激がlikingを抑制することが分かっている。要するに、快楽の感情のメカニズムは簡単ではない。
この快楽感情の複雑さを示す最たる例が「感情の鍵盤だ」だ。側坐核内の領域を丹念に刺激し、マウスの行動と対比させるマッピングを行うと、図4に示すようにlikingを高めるhot spotから嫌悪の反応を示すfear spotまでが、空間的に順序だって鍵盤のように並んでいることが明らかになった。この配置を彼らは「感情の鍵盤」と読んでいるが、快楽と恐怖が一つの領域に配置されることで、最終的感情を決めていることに驚嘆せざるを得ない。
図4側坐核の刺激実験により明らかになった「感情の鍵盤」
刺激により快楽を感じる部分から恐怖を感じる部分まで、順序良くスポットが配置されており、一つの領域でgo or not goの指示ができるように計画されている。
BerridgeとKringelbachの切り開いた快楽のメカニズム研究分野は現在急速に発展している。この結果、フロイトやスピノザが想像したように、快楽が私たちの行動を強く支配していることは間違いない。しかし、側坐核の複雑さからわかるように、これらの結果を人間の行動理解にどう当てはめていくかは簡単ではないが、hedonic spotとfear spotが常に共存して、私たちの感情を調節し、善か悪かを決めている現代脳科学の最先端に触れれば、スピノザも興奮するだろう。
またこのような感情に関わるスポットが存在する領域が脳内に散らばって、ネットワークされていることも明らかになった。このことは、このネットワークに、感情とは異なる様々な神経ネットワークが連結し、感情の強さや質を変化させ、また感情を行動に統合する。もちろんドーパミンにより作動されるWanting回路はまずこの感情回路と結合しなければならない。そしてこのようにして形成される「エス」は、さらに人間特有の脳回路と結合することで、超自我や抑圧された無意識が可能になる。この人間特有の回路とは何かを考えるときに基盤になるのが、この感情の回路であることは、現代脳科学が発達した今も、フロイトの時代から変わっていない。
次回からは人間特有の回路から生まれた最終的情報、言語について考えていく。
[ 西川 伸一 ]
ゲノムの発生
2016年8月1日
これから何回かにわけて、生物進化とその過程で生まれた情報について見ていく。この作業を通して生物進化を、様々な情報が生まれる過程として眺めてみたい。そしてこれらの情報を、「伝達性・コミュニケーション」と「環境の自己への同化」の観点から再検討することで、なぜ進化の究極に言語が誕生出来たのかに至る議論が出来ればと思っている。
前回述べたが、ある現象を特定のシグナルとして解釈できる生物機構がうまれると、この機構により解釈されることで、あらゆる現象は情報に変わる。この機構を理解することは当然生物進化の理解に欠かせないが、このシリーズでは、個体間、細胞間で伝達・コミュニケーション可能な情報に限って議論を進める。
元々、20世紀の情報理論は情報の伝達とコミュニケーションについての科学として生まれた。例えばシャノンの情報理論は、非物理的存在である情報が、伝達という観点から見ると、物理的物質(例えば電線)と量的な相互作用をすることができること、また情報が物理量による変化を受けてもコミュニケーションを成立させるための理論として始まった。また、選択された形質の子孫への「伝達性」はダーウィン進化論の中核をなす概念だ。生物にとっての情報は必ずしも個体間で伝達される必要はないが、今回は伝達・コミュニケーション可能な情報に限って議論する。
しかし、生物間で伝達とコミュニケーションに使える情報はゲノムだけではない。高等動物になると、ゲノム以外にも他の種の生物に伝えることのできる情報は数多く存在する。例えば言語も人間(生物)が持つ情報媒体だ。言語を獲得することで、私たちは自己を身体から解放し、ほぼ全ての人にほとんど無限の情報を伝えることができ、さらには他の個体と瞬時にコミュニケーションすることが可能になった。
このことを哲学者ヘーゲルはうまく表現しているので引用しよう。
「言葉は、自分を自分自身から分離する自己である。これは、純粋な自我=自我という形で、自らの対象となり、この対象態のうちにこの自己としての自らを支えるとともに、またそのまま他者と合流しており、他人の自己意識でもある。」(ヘーゲル 精神現象論:樫山欽四郎訳))
この様に伝達性・コミュニケーション性から見ると言語は情報進化の究極にあるが、言語に至るまで伝達可能な他の情報が進化の過程で現れた。これらの情報がどこに向かって、どのように伝えられるのかを見ていくのがこれからの作業だ。
情報の進化を理解するためのもう一つの観点に「環境の同化」を選んだのは、環境を同化しようとする指向性が進化の原動力と考えているからだ。環境による選択と言ってもいいのだが、ゲノム以外の情報が加わってくると、もっと積極的な環境の取り込みが起こっている。従って、ゲノム以降新しく生まれた様々な情報が環境を同化する過程を見ることで、進化の力学の本態を知ることができる。
環境の同化を考えることで、「進化は多様化した個体を環境が選択すると過程」といった紋切り型の議論を排して、進化の本当の力学を理解できるようになると期待している。そのため、個々の情報が進化にどう関わり得るかについても丁寧に検討してみたい。
伝達性を考えるとき、一つ注意してほしいのが、環境の変化を通して個体の活動が子孫に伝達できる点だ。あらゆる生物は様々な大きさの領域に拡がる環境を変化させる可能性を持っている。生物により変化させられた環境は、回り回って子孫の生存を制約できる。すなわち、生物による環境変化に関わった全ての情報は、こんどは環境からの制約という形で子孫に伝わる。少しわかりにくいと思うが、例えば言語情報を介する高次機能によりこれまでなかった新しい生活環境(例えば都市)が生まれたとしよう。こうして出来た都市は影響力のある環境として、子孫の生存や進化を制約する。これは人間だけの問題ではない。都市から排出されるスモッグで黒い羽を持つ蛾が都市では優勢になるという有名な「工業暗化」の話は、生物同士が環境を変化させることで、それぞれの進化に影響した例と言える。この生命の「歴史性」の問題は、進化を考える上でも重要な問題で、ゲノム以外の情報の進化への影響を扱う際常に留意する必要がある。
図1にこれから扱おうと思っている情報を図示したが、1)ゲノム、2)フェロモン、3)エピゲノム、4)神経回路と脳の高次機能、5)言語、そして最後に6)バーチャルリアリティーに限ろうと思っている。図に示すように、これらの情報を全て持つ生物は現在地球上には人間しか存在していない。また、地球上で生まれた伝達可能な情報は全て生命活動と連関しており、生命誕生と同時に生まれたゲノムから、約2万年前に誕生する言語まで、それぞれの情報の誕生は、生命の進化の方向性を大きく変化させ、また地球自体を作り変えてきた。
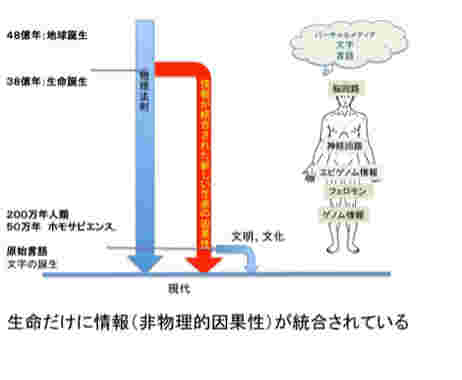
図1 進化から生まれた様々な情報:今回から順に扱おうとしている生命情報。これらは、38億年前のゲノム誕生と、約2万年前の言語誕生の間の生物進化過程で生まれた。
次回から図1に示した、生命情報、というより地球上に順番に現れた様々な情報進化として生物進化を眺め、それぞれの情報を伝達性と、環境同化性の観点から整理してみたいと思う。
ただ、地球に現れた最初の情報、ゲノムに関しては、これまで十分述べてきたので、今回手短に触れておくだけにする。
情報1:ゲノム
ゲノムを一言で定義せよと言われれば、私は「生殖を通して伝えることができる唯一の生物の情報」と定義している。生殖を通してだけ伝わるということは、ゲノムの伝達性は、ほぼ子孫に制限されていることを意味している。すなわち、複製を通して情報が子孫に共有され、それにより生きるためのメカニズムも共有される。
しかし他の生物の遺伝子が一部、あるいは丸ごと統合した名残を現在の生物にも見ることが出来る。例えばミトコンドリアやクロロフィル、あるいは水平伝搬でゲノムに入り込んだDNA断片だ。このため、生殖でしか伝えられないと言ってしまうのは言い過ぎとは思うが、高等動物になればなるほど「生殖を通してのみ伝えられる」という条件に従うようになる。
ただ忘れてならないのは、「生殖を通してのみ子孫に伝わる」というメカニズムと、他の個体の遺伝子を同化する過程を融合させた、「性生殖」という方法が生まれたことだ。
生物学的「性」とは、まさに個体間の遺伝子情報交換のことだ。これは原核生物にも存在し、一つの個体から他の個体に情報が移り、そこで相同組み替えが起こることで、異なる個体間の情報が一つの情報に作り直される。さらにこの方法でゲノム自体のコミュニケーション(相同組み換え)も可能になった。すなわちゲノムは伝達可能でコミュニケーション可能な情報になった。
このスッキリした定義は20世紀になり、少し危なっかしくなった。すなわち、人為的にゲノムを改変することが自由に行えるようになった結果、ゲノムの伝達性に関する制約条件は新たに変わろうとしているが、これについては言語を考えるときにもう一度議論したいと思う。
次に進化を環境の自己への取り込みという観点から見たとき、ゲノムが取りうる方法について見てみよう。
まず環境をRNAやDNAとして直接取り込むことが可能だ。この最たる例が、ミトコンドリアや葉緑体で、最初は体内の寄生生物として存在していたミトコンドリアの遺伝子が徐々に宿主のゲノムに移行して、宿主の自己に一体化していく過程は、まさに環境の取り込みとして見るとわかりやすい。
ミトコンドリアに限らず、環境 を情報として直接取り込む遺伝子伝搬は誕生初期の生物では普通に起こっていたと考えられる。ただ、この方法で直接環境からゲノムを取り込むことは、原始ゲノムから、単細胞、そして多細胞個体へと生物の自己の範囲が拡大するにつれてその頻度は下がっていった。特に多細胞動物になると、自己の制約から完全に独立した情報をそのまま取り込むことは、弊害の方が多くなったのだろう。この危険は、侵入した外来のゲノムの活性化を抑え込むエピジェネティックなメカニズムが進化しているのを見ると理解できる。
このことから、ゲノムは外来のDNA(情報)を容易に受け入れることができるが、自己という制約とは無関係に生まれた情報をそのまま取り込むことは、それがいかにゲノムの多様化に寄与するとしても、危険を伴うことがわかる。
例外として、既に述べたが性生殖という様式により、他のゲノムの断片を、別のゲノムに移すことが可能になっている。この場合、相同組み換えという機構が用いられるが、自己の制約の中で(相同性)ゲノム同士の情報が交換されるという様式が厳しく守られていると見ることが出来る。
もちろん、現在もなおトランスポゾンがゲノムの多様化に寄与する例はあるが、ゲノムが環境をとりこむための過程は、
を繰り返すことで進む。
繰り返すが、この稿では選択を環境の同化として捉えていく。誤解を恐れず言ってしまうと、これはキリンの長い首を、丈の高い木の幹という環境が同化した結果と考えることだ。
同じような同化の例として亀の甲羅の進化についての最近の研究を紹介して終わろう。論文のタイトルは「Fossorial origin turtle shell (かめの甲羅は穴掘りが起源)」で、7月号のCurrent Biology (Vol 26:p1887, 2016)に掲載されている。
亀の甲羅の進化は、通常頭などの体の柔らかい部分を隠す硬いシェルターとして進化したと考えられてきた。例えば強い歯や爪を持った天敵の存在が、心臓や肺を守る硬い肋骨の進化を促したとでも考えればいいのだろうか。すなわち天敵のいるという環境圧が硬い甲羅を選択したと考えられていた。
ところがこの論文では、最初からシェルターを肋骨から作ろうとすると、運動能力や呼吸機能が低下し、逆効果になる。一方、亀の甲羅は最初穴掘りに適した骨格として進化したと考えると、2億年から2億5千万年前に生息していたと考えられるカメが、強い肋骨に支えられた前腕、長い爪など穴掘りに適した骨格を持っていることとも話は合うと主張している。
この主張の真偽はともかく、シェルターの必要性の観点から見ると、先に甲羅を発達させるより、確かに穴を掘った方が早い。そして、その後カメだけはこのシェルターとしての穴を、甲羅という形で自己に同化することに成功したと考えると、まさに環境が自己に同化している。
進化での選択を、環境の同化と置き換えてみるのも悪くないと思っていただいただろうか。
さて、次回はまずフェロモンから始める。
[ 西川 伸一 ]
情報の進化:1 フェロモンから見る情報の伝わり方と意味
2016年8月15日
情報を伝えるというと、私たちはすぐ何らかの媒体で表現された情報を個体間でやりとりすることを考えるが、原始ゲノムから細胞へと自己を拡大したばかりの単細胞生物では、情報の伝達とは、個体と一緒に同じ情報が増殖することで、この結果、同じ情報を共有した異なる個体が生まれる。もちろんゲノムの複製はゲノムの多様化につながり、同じ情報というのは正確ではないが、ゲノムの伝達のためには複製以外の道はない。
こう言い切ると、「原核生物には水平遺伝子伝播で文字どおりDNAを媒体とした情報を伝えるメカニズムがあるではないか」と指摘を受けそうだ。しかし、ウイルスやプラスミドのような寄生体のDNAを別にすると、水平伝播で伝えられるDNA断片を情報と呼ぶことには問題がある。というのも、多くの場合DNAがゲノムに取り込まれても、取り込んだ方の個体がその情報を解釈する仕組みを持っていないことが多い。例えて言えば、私にとってロシア語の文章が情報になりえないのと同じだ。逆に、解釈できない異なる種由来のDNAを闇雲に取り込んでしまうと、情報が伝わるどころか、自らのゲノムの完全性(genomic integrity)が損われるほうが問題になる。事実、原核生物のゲノムは、環状化し、相同性のないDNAが挿入されないようにして、外来のDNAの侵入に備え、ゲノムのインテグリティーを守っている。
おそらくLUCA (Last Universal Common Ancestor:全生物の共通祖先)が誕生した最初の頃は、ゲノムがコードする分子の特異性も高くなく、少々の外来DNAの挿入は許容され、外来のDNAを積極活用する余地があったかもしれない。しかし、時間をかけて最適な機能分子が進化し、自己の範囲の中で(実際には細胞の中で)厳密な分子ネットワークが生まれると、変異や外来DNAの侵入は、進化には都合が良くても、個体にとってのリスクは大きい。
ゲノムのインテグリティーを守るためのDNA修復機構は、地球上の全ての生物に備わっており、LUCA誕生後おそらく急速に発展したはずだ。自らのゲノムと相同性を持つ場合にのみDNAをゲノム内に受け入れる仕組みは、まずゲノム修復を転用するシステムとして生まれ、その後DNA情報を交換する細胞の「性」の基本メカニズムとして発展する。
いずれにせよ、性を通して自分と相同性が高いDNA情報だけを伝える仕組みが発達し、ゲノム間の部分交換が可能になるまでは、他の個体からDNAが情報として伝えられることはなかったと言っていい。
ゲノムの新しい世代への伝達を生殖(増殖)に限ることにより、一個の個体由来の子孫がほぼ同じ分子を共有できることは重要だ。すなわち、細胞内外の同じメカニズムを全て共有する、自分のコピーが数多く周りに存在することで、様々な刺激に集団が同じように反応するポテンシャルが用意される。すなわち、生命を成り立たせている制約自体を集団へと増殖させることが可能になっている。
フェロモン
このような準備の後、新しく生まれた情報伝達機構が、フェロモンとエピジェネティック機構と言える。フェロモンは現存の原核生物や古細菌から存在しており、一方完全なエピジェネティック機構は真核生物から見られることを考えると、おそらくフェロモンの方が早く誕生したのだろう。
原核生物のフェロモンが最もよく研究されているのがQuorum sensing(クオラムセンシング)と呼ばれる過程だ。そこで、先ずこのQuorum sensingが何かから見ていこう。
Quorum Sensing
Quorumは日本語で定足数と訳される。すなわち、何かを行う決断するために必要な個体数のことだ。図1に示すように、Quorum sensingは個体濃度を常に測定して、個体数が一定数を超えた場合のみ、特定の分子発現(図では赤丸)のスウィッチをオンにする仕組みだ。
図1 Quorum Sensing 個体数を検出して(青丸の濃度)、一定の個体数に達したときだけ分子(赤丸)が発現する。このため定足数検出という名前がついた。(wikimedia
commonsより)
実はこの現象が最初に明らかになったのは、イカに住みついて光を発する発光バクテリアvibrio fisheriの研究からだ。すなわち、vibrio fisheriが蛍光物質を作るためには、菌がイカの体内で一定の濃度に達する必要があることがわかった。その後この仕組みが、様々な過程に使われていることが明らかになる。例えばQuorum sensingは細菌感染学でも重要な問題になっている。食中毒などのトキシンを分泌する事が出来る菌も、一定の菌数濃度に達しないと毒性を発揮しないことが知られている。このセンサーを破壊して、トキシンの分泌を止めることができると、有効な治療法になる。
Quorum sensingの分子メカニズムはほぼ完全に解明されている。このメカニズムには、定足数(菌濃度)を教えるための指標(インデックス)となる分子、その分子が一定濃度に達したことを感知する分子、そしてこの認識を様々な物質の生産につなげるための転写活性化メカニズムが必要になる。この分子機構については多くのバクテリアで解明が進んでいるので、ここではグラム陽性菌を例にQuorum sensingに関わる分子機構を示す。
図2:グラム陽性菌でのフェロモンによるシグナル伝達経路。説明は本文参照。
この系で定足数の指標になる分子がフェロモンだ。グラム陽性菌では私たちのペプチドホルモンと一緒で、先ず長めのペプチドが作られ、それが細胞外へ分泌されるとき短く剪定される。この仕組みにより、ペプチドを作った同じ細胞にセンサーが存在していても、一度細胞外へ出たペプチドだけが刺激できるようになっている。
一方、グラム陰性菌ではペプチドの代わりにラクトンを核にした有機化合物を使っている。これはエストロジェン(Estrogen:女性ホルモン)などの核内受容体に対するリガンドと似ていると言えるかもしれない。
Quorum sensingが働く菌は常にフェロモンを作り続けているが、菌濃度が低いと細胞外のフェロモン濃度は検出感度以下でとどまり、フェロモン受容体は活性化されない。ところが菌数が増えてくると、それに応じて細胞外のフェロモン濃度が上昇し、受容体は活性化され、標的遺伝子の転写のスウィッチが入る。これがQuorum sensingのメカニズムだ。
どちらのタイプのフェロモンにも2種類のセンサーが特定されている。一つは、細胞内に入ってきたフェロモンと結合して、転写活性のスウィッチが入る転写因子で、もう一つは細胞表面上でフェロモンと結合し、リン酸化反応を介してシグナルを伝える細胞表面受容体タイプだ。この構造を見ていると、人間がホルモンとして使っている、核内受容体(例えばエストロジェン受容体)と細胞外受容体(例えば成長ホルモン受容体)などと同じタイプのメカニズムがバクテリアから存在していることがわかる。
メカニズムはこのぐらいにして、フェロモンシステムの進化を考えてみよう。まず、原核生物で見られるフェロモンシグナル伝達システムの基本構造は、その後の生物で普遍的に見られることから、フェロモン自体の進化については割愛して、LUCAがフェロモンシステムを進化させてきた過程について考えてみよう。
理解を容易にするため、ここではフェロモンが細胞内で転写因子に結合して転写を活性化させる系だけを考える。ここで使われている分子メカニズムは、まさに細胞内で合成された分子を使ってフィードフォワードループを動かす系と同じで、あらゆる生物で普遍的に見られる。ジャコブとモノーが発見したトリプトファンに結合してトリプトファンオペロンを抑制するトリプトファンレプレッサーは最も有名な例だ。ただ細胞内部にメッセンジャーがとどまるフィードフォワード回路とは異なり、そのまま、或いはプロセッシングを受けたあとフェロモンとして細胞外に分泌されることで、自分だけでなく他の個体にも作用できるようになっている。
メッセンジャーを細胞内だけでなく、細胞外に分泌できるようにして他の個体にも情報を伝達する進化により、それまで全く単独で生きてきた生物は自己の範囲を広げ、他の個体と情報を共有し、同じ行動を取る一種の社会行動が可能になった。

図3 バイオフィルム(wikimedia commonsより)。バクテリアが一定濃度に達してEPSを分泌しフィルムを形成、それをバクテリアが一種の住処として利用している。
Quorum sensingの意義を考えるとき、バクテリアがEPSと呼ばれるポリマーを分泌して集合体を作るバイオフィルム形成過程で、同じ機構が働いていることは興味深い。バイオフィルムは単独では決して形成できない。形成に必要な細胞数に達しているかどうかの情報共有をQuorum sensingは可能にしている。すなわち、自己の範囲を拡大する進化をここに見ることができる。さらに、バイオフィルムは、環境の自己への同化が起こっている例として見ることができないだろうか?
ではこのシステムで生まれた情報の特徴について最後に見ておこう。まず情報の媒体にコード性はない。しかし、記号論的にいうとフェロモンは個体数のインデックスになっている。そしてこのインデックスを解釈する仕組みを共有することで、インデックス自体とは物理的に関係のない物質を作ることが可能になっている。このように、フェロモンシステムはゲノムと異なりコード性は全くないが、代わりにインデックス記号性を持ち、これを介して同時代の他の個体と情報の共有とコミュニケーションが可能になっている。
バイオフィルム形成と同じ情報共有システムはその後、あらゆる単細胞動物で発達を遂げ、多細胞動物への道を開く一つの条件となり、生物の新しい情報の重要な類型となった。
次回は、単細胞生物から多細胞生物への進化に必要だったもう一つの情報伝達・コミュニケーションシステム、エピジェネティック機構について考える。
[ 西川 伸一 ]
情報の進化II クロマチン構造を媒体とした情報
2016年9月1日
フェロモンの次に議論する情報は「クロマチン構造を媒体とした情報」だ。この情報は一般的にエピジェネティックスと呼ばれることが多い。ただ、エピジェネティックスという言葉の指す範囲は曖昧で広い。このため、ここではエピジェネティックスを「クロマチン構造を媒体とした情報」に限定する。
まず簡単に「クロマチン構造を媒体とした情報」、エピジェネティックスについて簡単に解説する。
私たち人間は40兆個近くの細胞を持っているが、全て一個の受精卵が発生過程で、発生プログラムに従って様々な細胞に分化した結果だ。リンパ球のような一部の例外を除くと、何百、何千種類もの異なる細胞ができるためにゲノムの変化は全く必要ない。細胞の形は異なり、作られる分子は全く違っていても、私たちの体の中の細胞は全て同じゲノムを持っている。このゲノムの変化なしに異なる性質の細胞が発生し維持されるメカニズムがエピジェネティックスだ(図1)。
図1 発生ではゲノムは変化せず、クロマチン構造の変化だけが起こる。従って、ほとんどの発生過程は、エピジェネティックな過程と見ることができる。(図はWikiを拝借)
次にこの発生の結果何が可能になったか考えてみよう。何百種類の細胞がただ発生したというわけではない。例えば皮膚の細胞は皮膚細胞を作り続け、血液細胞は血液を安定に作り続けることが可能になっている。同じゲノムを持っている細胞同士なのに、決して皮膚の細胞が血液に変わることはない。もしそんなことが一定の確率で起こったら私たちの体の統合性はたちどころに破綻する。
これを支えるメカニズムがクロマチン構造を媒体とした情報=エピジェネティックスで、原則として真核生物で初めて見ることができる。このゲノムとヒストンと呼ばれるタンパク質が互いに協調することで形成されるのが、クロマチン構造で、それぞれの細胞種でパターンが異なる。また、細胞分裂でヒストンが外れても、もう一度同じように再構成できることから、クロマチン構造を繰り返し形成し直すための一種の鋳型ができていると考えると判りやすい(図2)。
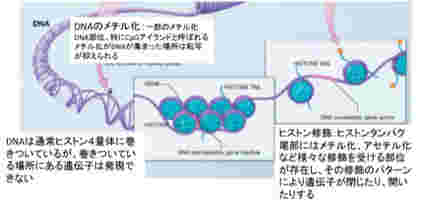
図2:クロマチン構造と修飾:ヒストンの少ないところはクロマチンが開いていると表現し、一般的にそこにある遺伝子は転写因子と相互作用する。一方、多くのヒストンが集まった部位はクロマチンが閉じていると表現し、ここにある遺伝子は転写因子から隔離されており、発現できない。(図はWiki
を拝借)
このクロマチン鋳型は、全ゲノムにわたって活性化部位と、不活性化部位を区別する印をつけることにより形成される。これらの印は、ノートの開きたい場所と、閉じておきたい場所につける付箋のようなものと考えるといい。
体の全ての細胞には、それが属する種類、すなわち皮膚細胞、血液細胞といった種類に対応する異なる鋳型が存在している。そして何よりも重要なのは、細胞分裂でこの鋳型も複製され、二つの娘細胞に伝わる。このような性質から考えれば、エピジェネティックスをクロマチン構造を媒体とした情報と呼ぶことは何の問題もない。また、この鋳型があるおかげで皮膚細胞は間違って血液細胞や脳細胞になる心配なく、皮膚細胞を作り続けることができる。実際、この鋳型を維持するメカニズムが狂うと、皮膚細胞が思いもかけない細胞へ分化するといった乱れが生ずる。ガンで見られる異型細胞の出現はこの例だ。
ではこのクロマチン鋳型とは何で、情報としてどのような役割を担っているのか見てみよう。すでに図2に図示したように、ゲノムは細胞の種類に応じてヒストンが詰まった閉じた状態と、結合しているヒストンが少ない開いた状態にわかれている。
この開いた状態、閉じた状態を決めているのが、染色体と結合しているヒストンのN末端にある領域に存在する数種類のリジン残基のメチル化とアセチル化、そしてDNA自体のメチル化だ。このゲノムの場所特異的なヒストン修飾パターンやメチル化DNAパターンが鋳型として子孫に伝わり、繰り返し働く。これを可能にするのが、発生過程で細胞分化に合わせて鋳型を書き込むメカニズムと、細胞分化のあと完成した鋳型を維持するメカニズムだ。
このメカニズムについての理解は最近急速に深まってきたとはいえ、今も研究途上の分野なので、ここでは解説は省く。ただ、多能性幹細胞を始め、様々な培養細胞のクロマチン鋳型を精密に調べる研究から、
などが明らかになった。
この結果は、発生や成長で細胞分化のシグナルに応じてクロマチン鋳型を書き換えるメカニズムと、書き換えたクロマチン鋳型を安定に維持するメカニズムが間違いなく存在することを示している。すなわち、塩基配列のようなコード性は存在しないが、クロマチンのパターンを解釈して複製できることから、エピジェネティックスをフェロモンの次に誕生した情報と呼ぶことができる。
最後にクロマチン構造を媒体とした情報の特徴を、ゲノム、フェロモンと比較しながら見てみよう。
前回見たように、フェロモンはリアルタイムの個体間のコミュニケーションを可能にした全く新しい情報で、ゲノムだけがどれほど複雑化してもなしえない可能性を拓いた。前回このフェロモンの役割について、単位あたりの個体数を感知して一定の反応を起こすquorum sensingを例に説明した。ただフェロモンが活躍するのはquorum sensingだけではない。研究が進むもう一つの領域が、バクテリア同士がプラスミドを受け渡すときに働くフェロモンだ。これによって薬剤耐性が多くのバクテリアに伝搬するため、医学にとっては重要な分野になっている。
図3:フェロモン刺激によりプラスミドを持つバクテリアから持たないバクテリアへのプラスミドの伝搬が起こる。このメカニズムで薬剤耐性遺伝子が伝播される例が多く知られており、医学でも問題になっている。
図3に示す腸球菌の系では、フェロモンはバクテリアのゲノムにコードされているが、その受容体は薬剤耐性を持つプラスミドにコードされている。この結果、フェロモンに反応するプラスミド(+)バクテリアと、フェロモンを分泌するプラスミド(ー)バクテリア同士が特異的な接合を起こし、プラスミド陰性のバクテリアにプラスミドを渡すことが可能になっている。実際には、プラスミド(+)バクテリアが自分のフェロモンに反応しないよう、うまく抑制システムができているが、詳細は省く。
この現象からわかるのは、フェロモンが引き金となって、DNAを媒体とする情報のやりとりが行われ、この結果、効果が一過性のフェロモンのシグナルが、遺伝子変化として記録されることだ。すなわち、一過性のフェロモンの効果を長期間保持するためには、バクテリアでは遺伝子の変化として記録する必要がある。しかし、プラスミドのような限られた情報の伝搬は簡単でも、ゲノムを書き換えたりすることは難しく、出芽酵母のような一部の例外を除いて、同じ書き換えを繰り返すことは難しい。
一方真核生物への進化により、一過性に誘導される遺伝子発現状態をもとに、クロマチン鋳型を形成する能力、すなわちエピジェネティック過程が誕生すると、遺伝子の書き換えなしに、外界からのシグナルを特定のクロマチン構造として書き残すことが可能になる。
これをバクテリアがゲノムに記録する場合と比べると、外来の情報を持続的に記録するという点では同じだが、
などの、ゲノムでは不可能とは言えなくとも、簡単ではない記録が可能になっている。
これがクロマチン構造を媒体とする情報の誕生により生まれた可能性で、この獲得には20億年、生物の進化全過程の半分以上の時間が費やされている。次回は、この新しい情報の進化と、この情報がもたらしたインパクトについて考える。
[ 西川 伸一 ]
外界の情報を記憶するためのエピジェネティックス
2016年9月27日
前回「クロマチン構造変化を媒介とする情報(<エピジェネティックス)」が、
1)情報の書き換えができる、2)様々な外界のシグナルの影響を比較的迅速に記憶できる、3)異なる個体間で同じ情報を共有して同じ反応を可能にする、の3点で、ゲノムを媒体とする情報と異なることについて述べた。今回はこの違いについてもうすこし具体的に掘り下げる。
これを読んでいる読者のほとんどは、ルイセンコの名前を聞いても誰かわからないだろう。ルイセンコは、メンデル遺伝理論だけでは説明がつかなかった植物の現象を捉えて、獲得形質は遺伝することを主張したソビエトの農学者だ。彼を政治的に支持したスターリン政府の後ろ盾を得て、論敵を逮捕・追放したことで、科学的議論に政治を介入させたとして科学史に大きな汚点を残した人物だ。彼の理論は我が国にも様々な影響を残したようだが、これは私が大学に入学するより前の話だ。幸い、ルイセンコと彼の理論をめぐる我が国での議論については本館名誉顧問の岡田節人先生が生命誌ジャーナルに紹介されているのでぜひ読んでほしい。
さてルイセンコがメンデル遺伝学の反証の根拠にしたのが「春化(春処理)」と言われる現象で、植物を低い温度に晒すことで開花時期が調節できるという現象だ。低温という環境で獲得した形質が、世代を超えて伝わる場合もあることから、獲得形質が遺伝すると考えたようだ。結局エピジェネティックス(遺伝した後)の問題で、ゲノム自体の変異を基盤とする遺伝学の問題でないことが明らかになる。このように、春化メカニズムは、エピジェネティックス、すなわちクロマチンの構造を介した情報の問題として現在解明が進んでいる。外界からのシグナルがクロマチン構造を媒体とする情報として書き換えられるメカニズムを理解するのにうってつけの例なので、まずこの春化とそのメカニズムについて見てみよう。
各々の植物は発生過程で開花を調節する機構を確立する。これには温度や日照時間を感知しながら様々な遺伝子の発現を調節する複雑なメカニズムが関与する。話をわかりやすくするために、実験のためのモデル植物として研究されているシロイヌナズナの開花を抑制するマスター遺伝子FLCに絞って春化の現象を解説する。
まず越冬一年草型シロイヌナズナのライフサイクルを見てみよう。秋に発芽、ロゼット型葉を形成してゆっくり成長しながら越冬する。その後、茎が伸びて開花、そして夏になると種ができる。この間の開花抑制遺伝子FLCの発現を見てみると、発芽時の秋には最も高いレベルにある。しかし、ロゼットが形成される冬には急速にそのレベルが低下し、春の開花期には0になる。開花が終わると夏に向けて急速に発現レベルが回復する。
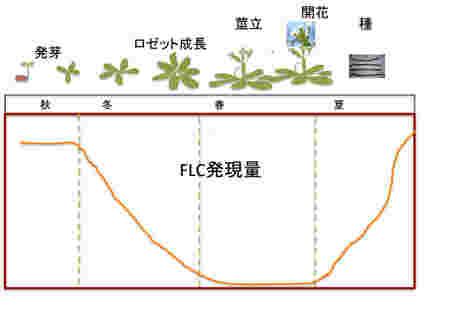
図1 シロイヌナズナの開花と、開花抑制遺伝子FLCの発現量。説明は本文参照。花と種の写真はWikipediaより。
FLCが開花抑制に関わることを頭において発現の季節パターンを見ると、開花の起こる春以外は開花抑制遺伝子であるFLCを発現させ、間違った時期に開花が進まないようできているのがわかる。すなわち、秋から冬にかけて気温が下がり始まるのを合図に、FLCの発現を安定に抑制するための準備を始め、この状態で急速に温度が上がると、開花を優先した植物の発生が進み、開花する。その後、種ができ、発芽するまでにFLCの発現を戻して開花を抑え、植物はエネルギーを成長に使うことが可能になっている。
次に、このパターンの背景にあるメカニズムを細かくみていこう。この季節に合わせた発現の調節にはFLC遺伝子領域とそこに結合しているヒストンのメチル化パターンが大きな役割を演じている。すなわちFLCの発現を抑えるときはこの領域に巻きついているヒストン3のN末27番目のリジンがメチル化されており(H3K27me3)、一方発現が高いときは4番目のリジンがメチル化されている(H3K4me3)(図2)。すなわち、クロマチンが閉じて転写が抑制されるか、あるいはクロマチンが開いて転写が行えるかは、ヒストンの標識の違いによって決まる。
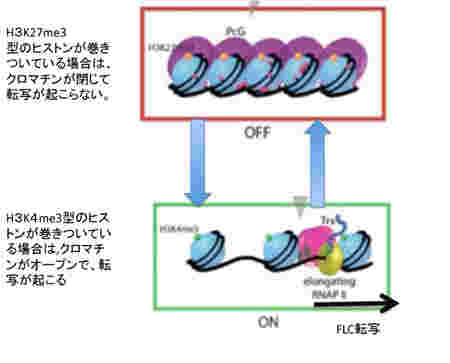
図2 ヒストンのメチル化によるクロマチンの構造のスウィッチ。図はES細胞分化時に起こるクロマチン構造変化についての説明をStem Bookサイトより転載。(http://www.stembook.org/node/585.html)
つまり、FLCに巻きついているヒストンのメチル化の異なる2つの状態を温度スウィッチでシフトできるようになっている。このようにクロマチン構造を媒体とする情報の誕生は、外界の変化を細胞自体の記憶に変えることを可能にした。
このヒストン標識によるクロマチン構造のスウィッチのメカニズムについてはずいぶんわかってきたが、複雑でその全像について解説するには本1冊必要になる。とりあえず、図にあるように、クロマチンの構造を維持する鍵になる分子が、ポリコム複合体とTrithorax複合体であることを理解しておけば十分だろう。
H3K4のメチル化と、H3K27のメチル化パターンを転換させクロマチン構造を変えることで大きな領域の遺伝子発現を調節するメカニズムは、実は酵母から人間まで保存されているメカニズムだ。事実、ポリコム複合体やTrithorax複合体という名前はショウジョウバエ突然変異の研究が由来となっている。例えば、ショウジョウバエから哺乳動物まで、体の前後軸に合わせて体節の特異性を決めているHox遺伝子が集まる大きなゲノム領域のどこまでを活性化し、どこまでを不活化するかは、まさにこのクロマチン構造変化により調節維持されている。
図3は哺乳類の発生でのHox遺伝子領域のヒストン標識パターンを図示しているが、Hox遺伝子の発現はクロマチン構造の変化で調節され、植物とほぼ同じメカニズムが使われていることがわかる(図3)
図3 クロマチン構造変化によるマウスHoxa1-13遺伝子の発現調節。内部細胞塊のような多能性幹細胞ではHox領域は完全にH3K27me3ヒストンで標識され、発現が抑えられている。前から後ろの体節に移行するのに合わせ(i)(ⅱ)(ⅲ)、Hoxa1からH3K4me3ヒストンは拡大していく。
出典(http://rstb.royalsocietypublishing.org/content/368/1620/20120367)
最後に、春化時のクロマチン構造の変化の際、他生物で見られない特徴について見ていこう。実は、FLC遺伝子発現を抑制するH3K27me3標識の範囲の量を調節できるようにできている。すなわち寒い時期が長いと、H3K27me3が結合するFLC遺伝子領域が拡大し、暖かくなってもこの抑制標識を外しにくくなる。これは、低温に長くさらされた植物の場合、開花が早くなり、開花期間は長くなり、さらに開花量も上昇するという量的な変化が、FLC遺伝子に結合するH3K27me3の量的な変化と相関していることを意味している。このように、外界からの情報を細胞内に量的な記憶として残すことができるのも、クロマチン構造を媒体とする情報の特徴だ。これをゲノム自体の調節機構で実現することはほぼ不可能だ。
春化を例に、クロマチン構造を媒体とする情報が、外界の情報を、細胞自体の情報に書き換えて記憶することを可能にしていることを示した。重要なことは、この記憶が書き換え可能で、量的にも変化させることが可能なことだ。進化が環境の自己への取り込みであるという観点から見ると、クロマチン構造を媒体とする情報は、ゲノムよりはるかに迅速に環境の自己への取り込みを可能にした新たな情報記憶の登場といえる。
今回例として使った春化では、環境の温度がクロマチン構造の情報として書き換えられる。一方、クロマチン構造の書き換えはフェロモンによっても誘導することができ、生命進化で誕生した新しい2種類の情報がここで出会う。すでに述べたように、フェロモンは個体間、細胞間のコミュニケーションに関わる情報と言える。一方、今回見てきたようにクロマチン構造を媒体とする情報は記憶媒体と言える。このように、コミュニケーションと記憶が揃った時、生命進化が大きく進む。こうして可能になったのが多細胞動物だ。次回はこの進化過程について考えてみる。
[ 西川 伸一 ]
コミュニケーション・伝達・記録
2016年10月3日
前回まで、ゲノム、フェロモン、クロマチン構造が媒介する情報と進化で生まれた新しい情報について見てきた。これらの情報を、伝達、コミュニケーション、記録という3つの情報機能について見直してみよう。
伝達、コミュニケーション、そして記録は情報が可能にする重要な機能だ。ゲノムはこれらすべての機能を併せ持っている。新しく生まれた情報には伝達機能は存在するが、フェロモンには記録という機能はなく、クロマチン構造を媒介とする情報はコミュニケーション機能を持たない。前回述べたが、フェロモンとクロマチン構造はゲノムの持つ情報を補完するように別々に誕生し、真核生物になると統合された情報システムへと発展する。そしてこの統合から生まれた新しい可能性が、多細胞動物の誕生の基盤になる。今回は、情報の記憶とコミュニケーション機能について整理をした上で、多細胞体制が生まれるまでに、生物の情報システムがどう発展したのか整理しよう。
まず、フェロモンやクロマチン情報が必要とされた背景、情報としてのゲノムの限界から見てみよう。
いうまでもなく、細胞が複製できるのは、ゲノムがDNAを記憶媒体として使って生物のオペレーションを指示する情報として働いているからだ。この記録機能と記録の伝達の原理については、DNAが2重螺旋構造をしていることが発見された時、解明されたと言っていい(図1)。
図1 2重螺旋の複製。2重螺旋が一本鎖にほどけ、それが2本の2重螺旋になることがDNAの構造から明らかになり、伝達と記録機能の原理が理解された。(wikipediaより)
ではコミュニケーション機能はどうだろう。複製されたゲノムは2つの娘細胞に伝わる。これは生殖を通しての情報伝搬だが、コミュニケーションではない。
ゲノム同士のコミュニケーションは基本的に相同組み換え機構を介して行われる。私たちの体細胞には、母親からの染色体と、父親からの染色体が存在するが、条件が整うと両方の染色体同士で相同組み換え(図2)が起こり、染色体の部分交換がおこる。これにより異なるゲノム同士のコミュニケーションが可能になる。
図2:相同組み換えを通した2つのゲノムのコミュニケーション(wikipediaより)
この機構はほぼすべての生物で保存されており、大腸菌から人間まで、異なる個体のゲノム同士のコミュニケーションにはこの機構が使われる。ただ、相同組み換えが起こるためには、DNAの切断が必要で、実際には放射線などによるDNA切断に対する修復機構として進化してきたと考えられる。通常の状態で相同組み換えが起こるためには、DNAを自然に切断する過程が必要で、細胞にとっては危険極まりない。このため多くの生物では、この機構を通したゲノム間のコミュニケーションが高い確率で起こる状況は限られている。例えば、人間では生殖細胞の減数分裂過程に限られ、体細胞で起こる頻度は低い。
このように、ゲノムの記憶、伝達、コミュニケーションの様式を見てみると、全てが生殖サイクルと完全に一体化していることがわかる。単細胞動物では、異なる個体同士が接合するとゲノムレベルのコミュニケーションが起こるのを観察できるが、コミュニケーションの範囲は1対1に限られており、一つのゲノム情報が同時に異なるゲノム情報とコミュニケーションすることはない(図3)
図3: ゲノムには情報が記録されており、複製を通して娘細胞に伝達される。また、接合、ゲノム間の相同組み換えにより、他の個体のゲノムとコミュニケーションできる。ただ、図からわかるようにゲノム情報の伝搬、コミュニケーションは全て一対一で個体(細胞)が対応しており、同時に複数の個体とコミュニケーションしたり伝達することはない。
ではなぜコミュニケーションが必要か、その利点を考えてみよう。ダーウィン進化を現代的に翻訳すると、様々な様式のゲノムレベルの多様化がまず起こり、ゲノムの多様化が形質の多様化として反影されると、この形質の環境適合性を指標に生殖優位性での差異が生じ、環境に適応した個体の比率が高まることで進化が進むと言えるだろう。このゲノムの多様化の一番の駆動力は、DNA複製に必ず伴うエラーだが、もう一段質の高い多様化の駆動力が相同組み換えによる多様化だ。
全くランダムな多様化と比べると、相同組み換えによる多様化は、ゲノムレベルのコミュニケーションを基盤にしている点が大きく異なる。通常これが起こるのは性生殖だが、この場合異なる環境で生きてきた個体がそれぞれ持っているゲノム同士の部分を交換し合う。視点を変えてこの過程を見てみると、その種が分離してから世代を重ねてきた異なる個体のゲノム同士、いわば環境によりすでに「テスト済み」のゲノム同士で情報交換が行われていることを意味する。それぞれの個体のゲノムがその個体が出会ってきた環境を同化していると考えると、この環境との相互作用の歴史が一つのゲノムの中へ再構成されているとみることができる。言葉を変えると、ゲノム間のコミュニケーションにより、より多くの環境との出会いが、ゲノムに同化される。これがゲノム間でコミュニケーションが行われることの重要な役割だ。
この様にゲノムには、情報としての全てが備わっている様に思える(事実それが進化の原動力となってきた)が、特定の生物集団の生存から見た時、ゲノムの対応能力には限界がある。すなわち、ゲノムだけで環境の変化に対応しようとすると、伝達やコミュニケーションが1:1に限られ、空間的にも、時間的にも急激な環境変化には対応できない。この限界を埋めるのが、フェロモンと、クロマチン情報だ。
詳しく検証したわけではないが、フェロモンは、細胞内の分子間相互作用ネットワークが、細胞外へ拡大したものと考えられる。フェロモンは接合の相手を惹きつけ、接合時の方向性(オスメスと考えて貰えばいい)を決める過程にしばしば登場する。これは1:1の個体間相互作用と言えるが、他にもクオラムセンシング(前々回)のように1:1の限界を超え、同時に多くの個体とコミュニケーションをとりあう手段を提供し、全ての個体が環境に対して同じように反応するための情報としても働いている。
この様に、生存環境に関する情報を他の個体と同時的に共有することを可能にしたのがフェロモンの機能だ。フェロモン自体の情報には記録機能がないため、フェロモンのシグナルは一時的な反応を誘導して終わるか、あるいはフェロモンの情報を引き金に、遺伝情報を書き換え(酵母の交配型の変換)や、プラスミドを介したゲノム情報のやりとり(前々回)を行うことで、経験が記録される。このゲノム自体の書き換えの代わりに、クロマチン情報の書き換え機構とフェロモンシグナルをリンクさせることもできる。実際、クロマチン情報の書き換えが、反応の多様性、伝達範囲の大きさ、伝達と記録の迅速性の点で、ゲノムの書き換えをはるかに凌駕しているため、これがその後の生物の主要な伝達様式になっていく。
前回みた様に、クロマチン構造を媒介とする情報は、書き換え、すなわち繰り返して記録が可能だ。このメカニズムが完成するのは、おそらく真核生物以降だが、これに必要な個々のメカニズムは原核生物や古細菌で進化してきた。事実DNAメチル化は原核生物から存在するし、ヒストンと相同性を持つタンパク質も古細菌から認められる。更には、遺伝子の相同性は欠如しているが、DNAと結合してヌクレオソーム構造を形成するためのタンパク質は原核生物にも存在している。
この様なヌクレオソーム構造が広く認められることは、プラスミドの様な小さなゲノムは別として、メガベース以上の大きさのゲノムは裸のままで存在することが不可能であることを示している。実際、バクテリアやアルケアでは、1mmの長さのDNAを1ミクロンの大きさの細胞内に安全に収納している。その上に、収納した全ゲノムをなんども複製することが求められる。ヒストンの様な、DNAをコンパクトに巻き取る仕組みなしには、ゲノムサイズの急速な増大はありえなかっただろう。
真核生物の誕生にはヒストン修飾を基盤とするクロマチン構造制御機構、分裂時の染色体分離に必要なチュブリン、そして形態や細胞分裂を制御できる細胞骨格分子アクチンが必要だが、この3種類のタンパクは同じ分子からできてきたと考える魅力的な仮説がある。すなわち、いずれもリン酸化やアセチル化による修飾を介した細胞の構造変化に関わる。この考えを採用した、真核細胞進化過程について図3にまとめてみた。
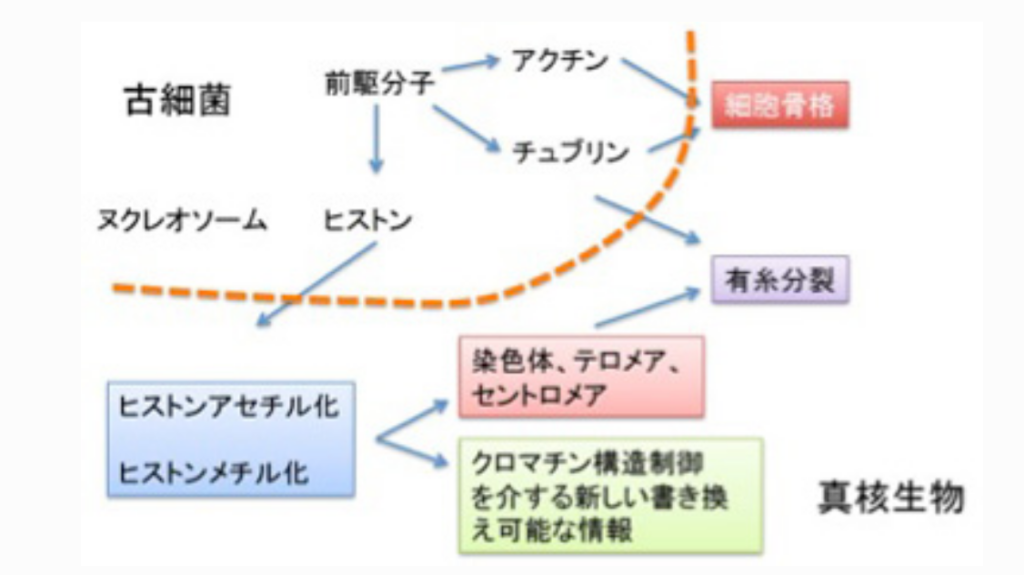
図3:古細菌から真核生物への進化をヒストンから眺めてみる。
ここでは話をヒストンに限るが、最初DNAをコンパクトに折りたたみ保護するヌクレオソーム形成分子として誕生したヒストンは、N末端に集まるリジン残基のアセチル化/脱アセチル化、メチル化/脱メチル化を介する修飾により、単純なヌクレオソームから、各ゲノム領域を構造的に標識するクロマチン構造へと発展した。これにより、テロメアやセントロメアのように、有糸分裂に必要な条件が整った染色体が生まれる。そしてゲノム領域を構造的に標識するヒストンの同じ能力が、遺伝子発現調節に使われるようになったのが、クロマチン情報だ。
春化の例でクロマチン情報は、環境のストレスを、多くの個体が同時に、同じシグナルとして感知し、このシグナルを同じ様に記録することを可能にしている。このようにクロマチン情報は複数の個体が同時に共有できる、ゲノムではなしえなかった情報と言える。
最初環境ストレスに対する反応として誕生したクロマチン情報の書き換え機構に、フェロモンによる細胞間情報共有機構が統合されると(図4)、それ自身では互いに未完成であった情報が補い合って、コミュニケーション、伝達、記録のすべて備わった新しい情報システムが完成する。
図4:環境ストレスによるクロマチン情報の書き換え機構にフェロモンが組み合わさると、より複雑な細胞間のコミュニケーションが可能になる。
図4で示した構造をよく見てもらうと、ある環境シグナルに対して直接対応する細胞と、その後フェロモンの作用で反応する細胞が別々に描かれているのに気づいてもらえたと思う。これは多細胞体制を想定して描いている。多細胞体制では、個体と環境、個々の細胞と環境、個々の細胞間、そして個体と個々の細胞間の4種類の関係が成立しており、この関係から生じる細胞の変化はすべてゲノムの変化を伴わない、クロマチン情報の変化が背景にある。例えば、個体と個々の細胞の関係には、同時的な情報の共有と、細胞レベルの経験の記録が必須で、これはクロマチン情報とフェロモン情報(細胞間情報伝達機構)だけで行われている。逆に多細胞体制成立から考えると、フェロモン情報とクロマチン構造を介する情報が誕生し、それが統合されることが、多細胞体制誕生の必須の条件であることになる。
[ 西川 伸一 ]
新しい情報と進化
2016年10月17日
前回まで、ゲノム、フェロモン、クロマチン構造が媒介する情報と進化で生まれた新しい情報について見てきた。これらの情報を、伝達、コミュニケーション、記録という3つの情報機能について見直してみよう。
進化についてのダーウィンの考えは、「集団は本来多様化する傾向を持っており、この多様化した個体の中から、環境に適合した個体が選択される」というものだ。ただ当時は全く理由の分からなかった個体の多様化の背景には、形質(フェノタイプ)よりはるかに大きな、ゲノムレベルでの多様化がある。
図1 ゲノムの多様化と自然選択だけから考える進化過程についての一つの表現。
ゲノム情報の多様化はエントロピーの増大と同じで物理的に自然におこる。この多様化したゲノムに対応する形質が環境にフィットした個体が選択されることになるが、これにより多様性(エントロピー)は、低下する。
この過程を単純な図式にすると図1のようになるが、この過程がただ続くとすると、せっかく獲得したゲノムの多様化も、選択により打ち消されるため生物が多様化する方向には進まないことになってしまう。
実際、全球氷結や隕石衝突など、生命自体にとって極端な条件が生まれると、ほとんどの種が絶滅し、生命の多様性は極端に低下する。ただ、通常はこのような条件が生まれるのは稀だ。なぜなら、地球は広く多様な環境が存在しているため、環境の違いだけ、それを同化した生物も多様化することになる。
ただこの説明のほとんどは、生命の情報としてゲノムしか想定していない。しかし私自身は、情報であれば全て進化に関わるものとして扱おうと考えて、これまで生命から生まれた全ての情報をリストする作業を進めてきた。もしフェロモン及びクロマチン構造を媒体とする情報のセットを生命の情報として認めるなら、当然この情報の多様化を統合して生物進化を考える必要がある。
そこでまず、フェロモンとクロマチン情報が新しい情報として加わることで、進化過程がどう変わるのか見てみよう。ダーウィン進化で問題になる情報は遺伝可能な情報であるという縛りがかかる。だとするとこの条件を満たす情報はフェロモンではなくクロマチン情報になり、クロマチン情報だけがダーウィン進化の駆動力として働く資格があるように見える。
しかし多様化できる情報をゲノムに限定しない新しい進化過程を構想するとき、フェロモンは記録情報でないからといって無視してもいいかというと、答えはノーだ。実際にはフェロモンにより新しい能力を獲得した集団が、環境を書き換えることで、能力の効果を記録してしまうことがありうる。例えば、古い旧家で何代もの子供が育ったとすると、先祖の建てた家という環境が、そこで育った子供たちに繰り返し制約を加える。この制約による変化も、ある意味で遺伝していると言って良い。
単細胞動物でも、多細胞動物でも、フェロモンは個体同士、細胞同士の行動の統合を可能にする仕組みだ。例えばクオラムセンシングを考えてみよう。この仕組みはフェロモンの濃度を感じて、周りに集る個体の濃度を感じ、十分な個体が集まったときに環境へ働きかけ、環境を自らの生存に適すように作り変えるのに使われている。このように、フェロモンに記録機能はなくても、環境側を変化させることで子孫へと一種の記録を残すことができる。この、環境を変化させるという記録の仕方は、後で神経細胞による情報を考えるとき重要になる(図2)。
他に、フェロモンは個体に働きかけ、接合を誘導したり、プラスミドを伝達したり、様々な方法で、ゲノムの多様化を促進する働きがある。こうして起こる多様化は、突然変異によるランダムな多様化ではなく、相同組み換えに基づくことで、多様化による生存への脅威を制約できるようになっている。すなわち、これまで生存を支えてきたゲノム情報の上に少しだけ変化を加える漸進的進化に適した変化の基盤になりうる。
一般的には、ダーウィン進化の駆動力、ゲノムの多様化は、DNA自体の化学的・生物学的性質により、自然に起こると考えられている。しかし、フェロモンが加わることで、変化させられた環境による新しい様式のゲノムや、クロマチン情報の選択が起こるようになる。また、接合誘導のような他の個体への働きかけの結果が、相同組み換えのような、単純なランダムなゲノムの多様化とは異なる、より制約を受けた様式による多様化が可能になる。このように、フェロモンだけでも、ゲノム多様化の様式を大きく変革できる(図2)。
図2 フェロモンの影響は様々な形で記録できる。説明は文中
次に、記録できる新しい媒体クロマチン情報は、それ以前の進化過程をどのように変化させただろうか?
前回述べたように、ヒストン修飾やDNAのメチル化を介したクロマチン情報は、テロメアやセントロメアの形成を通してゲノムを構造化することに成功した。これにより、環状DNAでは限界があったゲノムサイズの増大が可能になる。そして、同じクロマチン構造変化を誘導するメカニズムが遺伝子転写のOn/Offを指示する情報へと発展する。このクロマチン情報によって、遺伝子発現パターンの調節、記録、そして子孫への伝達が可能になっている。
クロマチン情報で常に問題になるのが、「エピジェネティックな記録は子孫に伝わるのか?」という遺伝の問題だ。しかし、多くの細胞は分裂を繰り返しても、同じクロマチン情報を維持できる機構を有しており、原理的にはクロマチン情報は遺伝できると考えていい。
もちろん多くの動物の初期発生過程では、卵子と精子などの配偶子が持っていたクロマチン情報が一旦キャンセルされたあと、体細胞とともに生殖細胞が新たに発生する。従って、新しい個体が発生する過程では、親の持っていたクロマチン情報は消えることが多い。ただ、iPSリプログラムによるクロマチン情報の変化についての研究からわかったように、クロマチン情報を完全にリセットできないケースがある。このような場合、エピジェネティックな状態が子孫に伝わったとして、大きく取り上げられることになる。
いずれにせよ、単細胞生物や、あるいは培養細胞のクロマチン情報を調べた研究から、細胞分裂を経てもクロマチン情報は維持され、子孫に伝達されるのが普通だ。したがって、単細胞動物の進化過程では、クロマチン情報も子孫に伝達される情報として、進化過程の駆動力として貢献できたはずだ。
次にクロマチン情報が加わると、どう進化は変わりうるだろう?
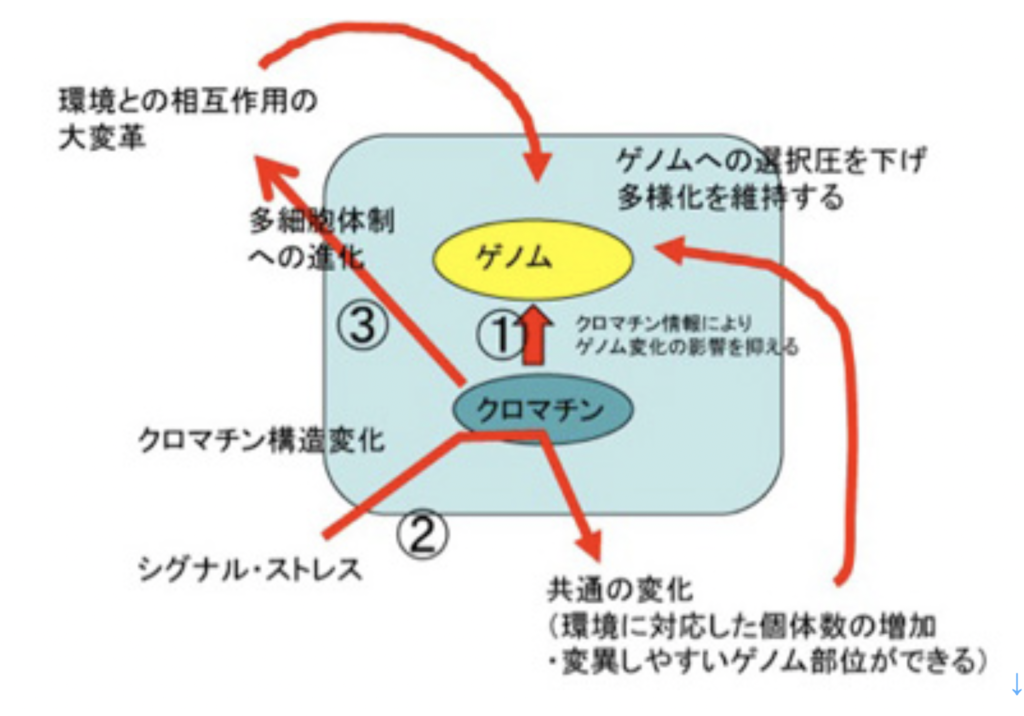
クロマチン情報の誕生による、進化様式の変化。説明は文中。
以上、クロマチン情報とフェロモンが情報に加わることで、進化の様式が大きく変革することを理解してもらえたのではないだろうか(図3)。
これまで、進化の様式が大きく変化するためには、選択圧が高まることの必要性が強調されることが多かった。しかし、情報を統合的に考えることで、私は逆に淘汰圧が下がり、最終的に種の多様化を決めるゲノムの多様化が保障されることが、生命の多様化には重要でないかと思っている。これまで述べてきたように、ダーウィン進化の結果は環境の自己への同化だ。多くの環境要因を同化できるということは、種が多様化することと同じだ。図1に示したゲノムしか情報がない場合の単純な進化過程をいくら積み重ねても、真核生物誕生後の急速な進化を説明することはできないのではないだろうか。ただ残念ながら、これは私の個人的意見に過ぎないと断っておく。
ゲノム、フェロモンからクロマチン情報へと進んだ情報の多様化と統合過程の繰り返しが、次は神経細胞の誕生から始まる。
[ 西川 伸一 ]
動物と神経の誕生
2016年11月1日
神経系がゲノムやクロマチン情報とは全く異なるレベルの情報システムを形成していることは明らかだ。ただ、これは神経系が高度に発達した動物についての話で、神経細胞誕生=新しい情報の誕生ではない。即ち、無生物から生物が誕生した過程と同じ問題が、神経系から新しい情報が生まれる過程に存在している。そこで、今回から神経細胞の誕生によって可能になった情報システムの成立について、動物進化過程を振り返りながら考えようと思っている。
この新しい情報システムは、もちろんゲノムやクロマチン情報とは密接な相互依存関係にあるものの、半独立の情報システムを形成していると言える。
例をあげて説明しよう。これまで述べてきたフェロモン、クロマチン情報は、ゲノム情報から独立して情報として働くことはできない。例えば、クロマチン情報はゲノムを構造化するとともに、この構造を変化させることでゲノム上の様々な大きさの領域にある遺伝子が利用可能かどうかを決定する(65話図3)。すなわち、クロマチン情報はゲノムとの関係を前提に複製可能で、その情報はゲノム上の遺伝子発現の違いとして表現される。誤解を恐れず例えるなら、ゲノム上の遺伝子の発現を指令するアルゴリズムと言える。
同様にフェロモンも、クロマチン情報やゲノム情報の変化を誘導することが主要な機能だ。フェロモンやクロマチン情報は独自の情報として振舞っても、ゲノムから決して独立することはない。
しかし、神経系による情報の解釈、生成、伝達は、ゲノムは言うに及ばず、クロマチン情報からもかなりの程度独立していると考えられている。実際、人間の脳が生み出した情報は、ゲノムやクロマチン情報からは独立している。だからこそ、神経系をコンピュータで再現できると考える科学者は多いし、また脳自体をコンピュータの一種として理解しようとする科学者は多い。
私自身は、神経を媒体とする情報システムは、コンピュータと同じで、神経細胞同士が階層的なネットワークや回路を基盤にしたシステムだが、その回路自体がクロマチン情報やゲノム情報と相互作用できることが、神経系独自の情報システム形成に重要な役割を果たしていると思っている。ただ、この問題については最初から議論するのを避けて、まず神経細胞の誕生から脳の誕生に至る神経系の進化を段階的に見ていきながら、それぞれの段階で神経系を情報の観点から眺め直すとともに、結合性とクロマチン情報や、ゲノムなどの下位の情報との相互作用について考える。
最初は神経細胞の誕生だ。現存の生物の系統樹から判断すると、神経細胞は動物、すなわち動く能力を持った多細胞生物の誕生とともに生まれてきたことは間違いない。図1に示した系統樹では現存の多細胞生物を5種類に大別しているが、海綿動物とセンモウヒラムシには、いわゆる神経細胞は存在しないことがわかっているが、残りの動物は全て神経細胞を持っている。
図1:動物の系統樹 現存動物の系統樹。この図では、神経細胞や筋肉細胞を持たない海綿やセンモウヒラムシを、他の動物から区別して表しているが、ゲノムから得られる系統樹とは一致しない。
では神経細胞は動物が進化した後現れたのか?
海綿やヒラムシに最初から神経がなかったかどうかは議論が分かれている。というのも、図1のクシクラゲは、ゲノム系統樹から見ると他の4種の動物から最も早く分離した系統で、左右相称動物から見て系統上最も離れている。ところがこのクシクラゲに神経細胞が存在することから、より左右相称動物に近いヒラムシや海綿にも最初は神経細胞が存在し、その後神経細胞を退化させたと考える説もある。
これとは逆に、神経細胞と、他の興奮性の細胞とを区別する指標として用いられるナトリウムチャンネルは左右相称動物だけにしか見つからないことから、クシクラゲと他の動物が別れた後、クシクラゲと左右相称動物は独自に神経細胞を発生させたと考える説もある。
いずれにせよ、最初の神経は私たちが一般的に持っている軸索、細胞体、樹状突起からなる典型的な神経ではなく、外界からの刺激に反応し、その興奮を他の細胞に伝達する能力を備えていても、より普通の細胞に近い形態を持っていたのではないだろうか。実際、興奮性の細胞系列の一部は、興奮を力に変える筋肉細胞へと発展する。一方、様々な刺激を感知して、他の細胞に伝達する能力を持つ細胞から神経細胞が生まれたのだろう。細胞の興奮に必要なイオン勾配の維持機能、そのイオンを選択的に通過させ膜電位を発生させるイオンチャンネル、そして興奮を他の細胞へ伝える化学システムは動物の誕生前に進化し、採用を待つだけになっていた。
では、神経細胞が生まれることで何が可能になったのか、現存の生物で見てみよう。
図2にゴカイの幼生の光受容体神経細胞について示すが、環形動物幼生の神経系は、まだ組織化されない単独の神経細胞が、どう神経細胞進化の初期に使われていたかを考えるいい材料になる(図2)。
図2:ゴカイ幼生の走化性
G.Jekely et al : An option space for early neural evolution,
Pil.Trans.R.Soc.B :
370:2015.0181 Fig2Aより改変、この図の元となったoriginal 論文はJekely et al, Nature 456, 395, 2008)
ゴカイの幼生は繊毛を使って水中を浮遊しているが、光を感じると繊毛運動を調整して光の方向へ移動する走光性を示す。これに関わるのが図2で示した3種類の細胞、色素細胞、神経細胞(光受容体細胞)、そして繊毛を持つ上皮細胞だ(original 論文はJekely et al, Nature 456, 395, 2008)。図に示す様に色素細胞とそれに結合する神経細胞により色素細胞で吸収された光エネルギーが神経興奮として受容され、同じ神経細胞が繊毛上皮とコリン作動性のコンタクトを形成することで繊毛の動きを調節している。片方の目にだけ光を当てると、光を受けた方だけ繊毛の運動頻度が低下し、結果幼生は光の方に進む。
この研究から、神経細胞の出現により、外界の物理的刺激を感知して、他の細胞(近くの繊毛上皮細胞)にそれを伝達して、活性を変化させる一連の過程が可能になっているのがわかる。
光や温度、あるいは圧力などの物理変化に素早く対応することは、生物の生存にとって重要な条件だが、神経細胞出現後の素早い反応はフェロモンのような化学的シグナル分子だけでは実現できない。このため、光などの外界の物理的刺激に素早く反応するための分子メカニズムは動物誕生以前から存在していた。例えば、単細胞生物クラミドモナスの走化性に利用される光を感じて開くイオンチャンネル、チャンネルロドプシンはその典型で、この分子を使った光遺伝学は、脳研究を席巻する勢いだ。
このような様々な物理刺激を感知して素早く個体の行動を変化させるシステムは、高等動物への進化過程で神経系の属性へと集約していくが、ゴカイの幼生の走光性行動には様々なメカニズムが組み合わさっている。
動物の運動のほとんどは筋肉、あるいは筋上皮と呼ばれる筋肉と上皮の中間のような細胞により行われる。実際、ともに興奮性の細胞と言える神経と筋肉は動物の誕生とほぼ同時に出現している。一方繊毛による運動は例えばゾウリムシなどの繊毛虫から見られるシステムで、より古い起源の運動生成システムと言える。私たち人間も含め、繊毛運動が神経支配を受けることは珍しく、その方向性やパターンは上皮の持つ極性を含むplanar polarityと称される特性により決められている。ゴカイの幼生は、この古いplanar polarity様式と神経支配という新しい様式を組み合わせて運動を調節している面白い例だと言える。
神経細胞により繊毛運動の強弱を直接神経支配することは難しい。結局ゴカイの幼生で見られるように、繊毛を持つ上皮自体に働いて、その細胞の持つ繊毛の運動を全て止めるという方法をとらざるをえない。
またもともと繊毛は早い大きな動きには向いていない。このため動物では、神経とアクトミオシン収縮系を備えた筋肉とがセットで働く”運動”が優勢になっていく。この組み合わせにより、一つの神経シグナルを複数の筋肉細胞の収縮に使うことが可能になり、大きな力を発生させることができるようになった。
もちろん、神経支配による上皮の活性調節は、ホルモン分泌の神経支配などとして今も見ることができる。
以上繰り返しになるが、ゴカイの幼生で見られる神経細胞の役割は、1)物理刺激を感受して、2)それを素早く他の細胞に伝達し、3)個体全体を刺激に対応させる、ことに尽きる。また同じ系が収縮性の筋肉細胞と組み合わさると、繊毛運動をはるかに超える大きな運動が可能になる。
最後に、神経細胞誕生により生まれた新しい可能性を情報と進化の観点から考えてみよう。
高等動物の脳神経システムには、間違いなく新しい情報システムが存在しているが、神経細胞や筋肉細胞の誕生により、これにつながる新しい情報システムが誕生したとは言えない。例えばクオラムセンシングでのフェロモンの働きに見られるのと同じ、環境の変化を感受して決まったアクションを起こす、インプット・アウトプットの関係が、より複雑化しただけと言っていい。事実、ナトリウムチャンネルの出現といった、新しい分子の誕生によるメカニズムの多様化がこの過程で起こったものの、例えば脊椎動物進化で抗体分子が現れるといった、本当のイノベーションは起こっていない。
しかし、物理刺激を感知できるようになり多様な環境変化に対応できるようになったこと、素早い反応が可能になったこと、そして筋肉の誕生により大きな運動が可能になったことは、環境を自己に同化する進化の駆動力の観点から見ると、大きなイノベーションだったと思える。
環境を感知して自己を守る反応という点ではフェロモンと同じだが、素早く、方向性を持った動きが可能になると、好ましくない環境を回避することで生存する個体数は当然増える。さらに、筋肉が生まれると素早い大きな回避行動をより刺激に合わせてコントロールすることで、多様な生息環境を選ぶことができる。これは環境を作り直したのと同じことで、環境の自己や種への同化が起こっていると見ることができる。こうしてより適した環境に移動することで、当然選択圧は下がり、多様性を維持する方向に働く。
このように、フェロモンも、神経や筋肉も、ゲノム情報による自然選択をできるだけ回避する方向に働く。実際、生きている個体の適応性は、それ以前のゲノム多様化と、選択の結果だ。新しい現在の環境に適合するかは試されたことがない。これに対し、新しく生まれた情報システムは全て、今生きている個体の生存に関わる。このように、今という時間への適応を拡大することが、生物の多様化に貢献したと思う。神経や筋肉で言えば、運動性が質及び量の面で大きく高まることで、動物は急速に地球上の様々な環境へと拡大できた。すなわち、過去の進化で獲得された環境適合性に合うよう個体が環境を選び、場合によっては改変することで、選択をかわしてゲノムをさらに多様化できたと思う。
新しい情報誕生まではまだまだだが、神経細胞の誕生はさらに新しい展開を見せる。次は記憶の誕生について見ていく。
[ 西川 伸一 ]
神経記憶 I
2016年11月15日
今回から記憶について考える。
記憶自体は情報ではない、しかし、皆さんのパソコンでの情報処理に記憶素子が欠かせないように、情報が成立するために欠かすことができない機能だ。情報を論じるために記憶の問題を避けて通れないことを理解してもらうため、これまで見てきた情報を、記憶という観点から見直すところから始める。
通常「記憶」という言葉は、経験によって脳内に誘導された神経活動パターンを、時間を経た後で呼び起こすことを意味する言葉として使っている。この神経記憶については後に考えるとして、この言葉が私たちの神的活動にとどまらず、あるパターンを繰り返し呼び起こす現象を表現するのに使われていることを思い出して欲しい。例えば、形状記憶合金というと、形が壊れてもまた自然に同じ形を取り戻すことができる合金を意味している。
生物学でも記憶という言葉が使われる現象は多い。例えば免疫記憶は、一度経験した抗原に対して免疫系が最初よりより速やかに、しかもより強い反応を起こす過程と定義できる。免疫記憶の背景には、かなり複雑な細胞間相互作用が存在するが、極めて単純化して説明すると、経験した抗原に反応するリンパ球が選択的に増加する、あるいは寿命が長くなることにより記憶が形成される。すなわち、経験を、この抗原特異的リンパ球の数や活性の変化に変換して記録している。
このように何らかの形で一定のパターンを記録することを記憶と考えると、生物にとって最も根源的な記録を担うDNAも一種の記憶と呼ぶことができる。実際、ゲノムは、ゲノム自身の複製だけでなく、それを持つ生物の複製も可能にしている。そして、複製とは同じパターンが繰り返すことに他ならない。もし、コンピュータのメモリーのように記憶=記録情報だとすると、ゲノムをメモリーと呼んでよさそうに思える。
しかし、塩基配列としてゲノムに書かれている情報を決して記憶と呼ぶことはできない。なぜなら、ゲノム複製も、細胞複製も、ゲノム情報だけでは実現できず、ゲノムの情報に基づいてオペレーションを行うための様々な条件が揃って初めて可能になる。以前、マイコプラズマの再構成実験を例に(http://www.brh.co.jp/communication/shinka/2016/post_000013.html)この問題について議論したが、今の所ゲノムは生きている生物にしか移植できない。同じことは、発生初期の受精卵では胚自体のゲノムからの転写がほとんどないことからもわかる。個体の一生を通じてあらゆる細胞で働くことができるゲノムも、まず母親から受け継いだRNAから作られた分子による発生のプログラムが活性化されて初めて、その個体の情報として働き始める。
このように、記憶は記録された情報ではなく、体験、記録、呼び起しという順序で進む一連の過程全体に他ならない。例えば、図1に示すように、ゲノムは情報として二つの役割を担っている。
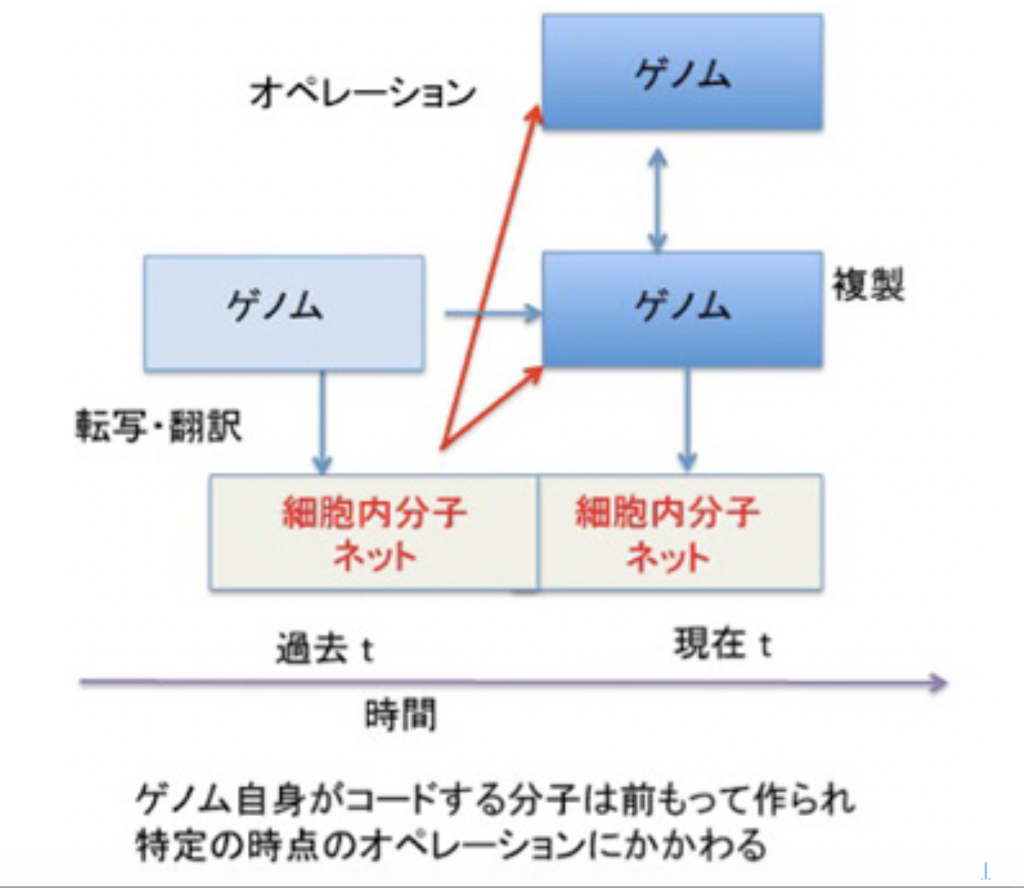
図1 ゲノムは生物の持つ最も基本的記憶過程と考えられる。説明は文中
一つは自分を複製するための情報と、複製のための鋳型としての役目だ。ただ、ある特定の時点(図1:現在 t)のゲノムのオペレーションは、同じゲノムによりその時点より前に作られた(過去 t)分子ネットワークによりオペレートされている。生命特有の鶏が先か、卵が先かという問題がここでも現れる。いくら外来遺伝子を細胞の中で機能させられるからといって、ゲノムとそのオペレーションは互いに分離不可能で、ゲノム=記録として切り離すことはできない。
ではなぜ今使っているコンピュータでは、アプリケーションや記録をメモリーに記録して分離することができるのか。これは、コンピュータ自体が一定の命令で動くように人間により作られているからというしかない。生命に、コンピュータにとっての人間のように、明確なデザインの出し手は存在しない。
ゲノムのオペレーションがそれ以前に準備された分子ネットワークに依存しているとすると、ゲノムのオペレーションに関わる分子ネットワークを辿れば、結局最初の生命誕生時にゲノムをオペレーションした分子ネットワークに行き着く。すなわち、ゲノムが38億年前に誕生した最初のゲノムまで連続しているのと同じで、細胞内の分子ネットワークも途切れることなく過去から現在そして未来と、ゲノムとともに受け継がれる。このように、現存の生物の記憶には無限の過去の経験が持ち込まれていることになる。こう考えると、進化自体DNAを記録媒体とした記憶と考えることができる。言いかえよう。ゲノムを記憶という観点から考えると、必然的に38億年という全進化史が含まれる壮大な過程と見ることができる。どの体験がゲノムに記憶され現存の種に維持されているのかを研究するは、進化学に他ならない。
一方、クロマチン情報は、基本的には各生物の一生という時間枠の中での記憶と言える。この意味で、神経記憶と同じだが、クロマチン情報が記憶と定義することに抵抗を感じる人も多いだろう。ただ思い出して欲しい。生物発生では、卵から個体誕生まで、ほぼ同じ過程が概ね正確に繰り返される。この過程では、ゲノムは変化せず、時間、場所に応じてクロマチン情報が変化していく。例えば発生学でよく研究される両生類の初期発生を例に見てみると、どの場所に精子が入るかは偶然の結果だが、侵入点が決まると体の大まかな体制が決まるよう、すでに卵細胞のゲノム、クロマチン情報、分子ネットワークが準備されている。
核移植クローン実験から明らかになったように、卵細胞質の分子ネットワークの多能性プログラムを維持する能力は高い。しかし、この分子ネットワークは卵割に伴い内外から新しい刺激を受けて分子ネットワークを変化させると共に、次々と生まれてくる細胞のクロマチン情報を変化させて、分化可能性が制限された分化細胞を形成する。一旦分化細胞のクロマチン情報が確立すると、分化した細胞の性質は安定に維持される。
クローニングやiPSにより示されたように、分化した細胞のクロマチン情報による分化制限を元に戻すことはできないわけではないが、普通はこの制限のおかげで、安定に同じ運命の細胞を作ることができる。こうしてそれぞれの細胞でクロマチン情報を中心に新しい記憶システムが成立する。
このクロマチン情報を中心とする記憶システムは、プログラム前の分子ネットワークと、外から入ってくる新たなシグナルにより少しづつ変化するが、このオペレーションメカニズムについては研究が始まったばかりでわかっていないことが多い。ただ最近、比較的少数の細胞でゲノム全体をカバーするクロマチン情報を調べる手法が開発されているので、急速な研究発展が期待できる。
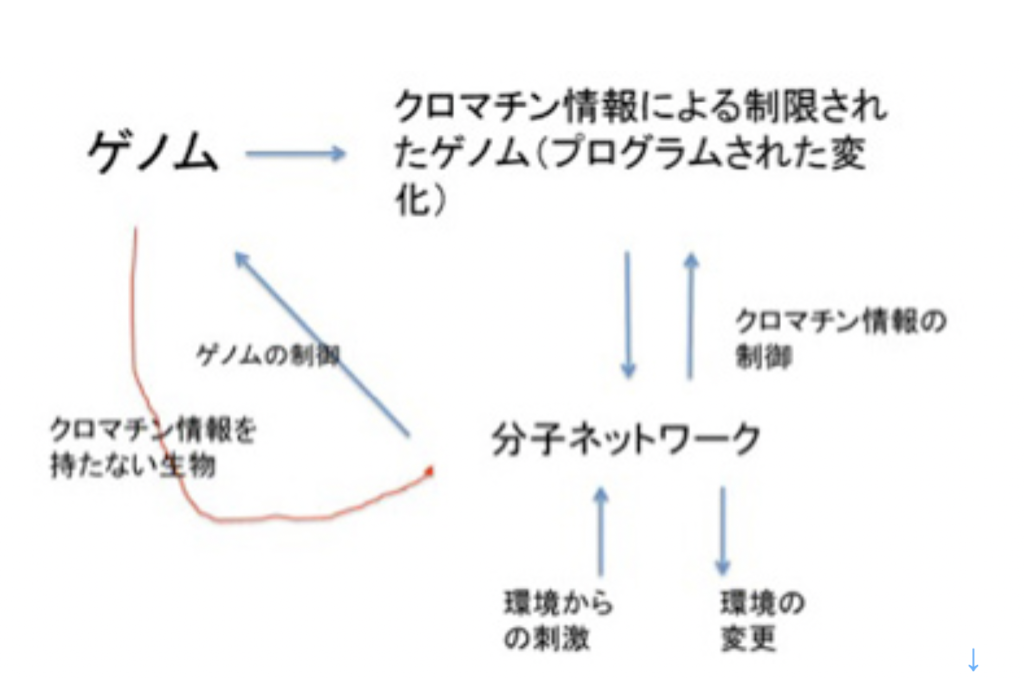
図2:クロマチン情報を中心とする記憶システム。説明文中
ゲノム、クロマチン情報、分子ネットワーク、外界からの刺激などを記憶の観点から見た関係を図2に示す。ここでは、ヒストン修復と、DNAのメチル化の制御によりクロマチン情報が書き込まれることで、特定の細胞を繰り返し作るための長期記憶が形成される。この記憶=クロマチン構造の維持と変更システム、により制限を受けた転写により生まれる分子ネットワークが、分化した細胞のオペレーションの基盤になる。各細胞の分子ネットワークは内外からの(周りの細胞も含む)様々な刺激により変化する。この変化は、一種の短期記憶と見ることができるが、クロマチン情報の制限を外せない限り、長期記憶にはなりえない。ところがある特定の刺激が分子ネットワーク内に揃うと、ヒストン修飾やDNAメチル化の変化による、クロマチン情報の書き換えが起こる。これにより、細胞の分子ネットワークの変化として短期的に維持されていた記憶が、より長い期間維持される長期記憶に変化する。
ある特定の細胞のクロマチン情報の書き換えを可能にする分子ネットワークの変化について研究しているのが、リプログラム研究で、山中4因子の発見に端を発して、様々なリプログラミング、すなわちクロマチン情報書き換えのための条件が明らかになりつつある。
神経記憶から脱線したが、図1、2に示した記憶システムは、真核生物のあらゆる細胞で見られる。従って、神経細胞記憶もこの2つの記憶システムの上に発展したといえるが、これまでの記憶システムとは大きく違っている点がある。すなわち神経記憶は、動物の誕生とともに新しく生まれた興奮性の神経細胞同士の結合により(ここでは、神経細胞を、同じ前駆細胞から生まれるニューロン、アストロサイト、オリゴデンドロサイトを全て含む広い定義を用いる)新たな記憶システムを成立させている点だ。
この新たな神経記憶の特徴については次回から考えるが、最後に神経記憶もクロマチン情報を中心とする記憶システムの上に構築されていることを、前回に例として使ったゴカイの幼虫の神経系をモデルに見ておく。このモデルを選んだのは、これが一本の神経細胞だけで形成されている最も単純な神経系で、なおかつ神経細胞の基本的特徴を備えているからだ。そこで、ちょっと想像をたくましくして、この系での記憶について考えてみた。もちろんこれから述べることは証拠があるわけではなく、神経記憶自体は回路形成なしでも考えられることを知ってもらうために考えたたとえ話だ。
さて前回述べたように、ゴカイ幼虫には、光をトラップする色素細胞と協調して光に反応し、その興奮を繊毛細胞に伝え、繊毛の動きを止める神経細胞がある(前回図2、および今回図3)。この神経は感覚神経と運動神経の両方の役割を持つため、神経回路は形成しない。
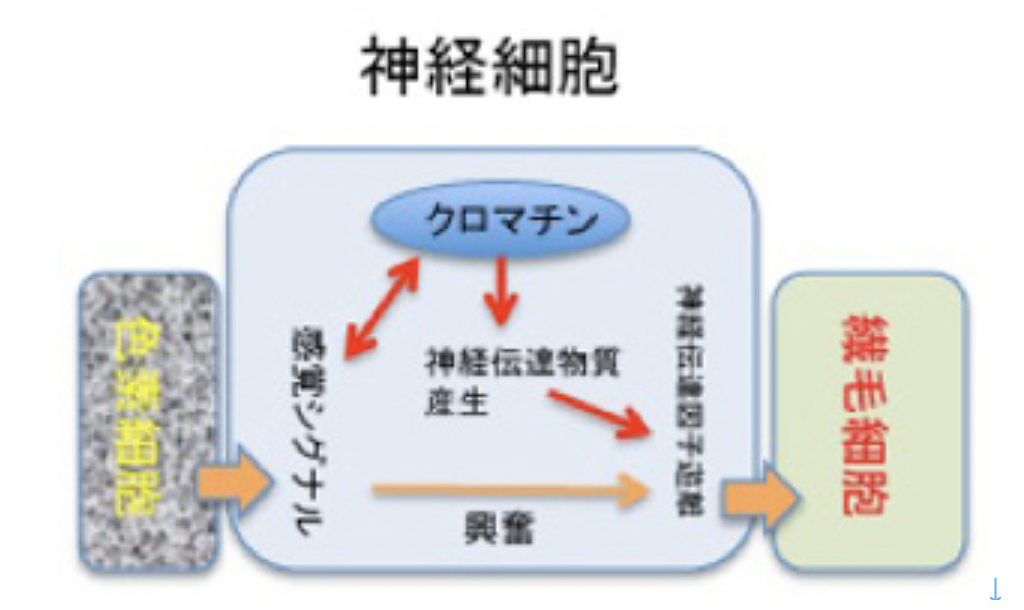
図3:ゴカイ幼生の感覚運動神経システム 説明文中
この系では神経細胞が感覚刺激を感受すると、チャンネルが開き細胞が興奮、その興奮によりアセチルコリンが遊離、繊毛細胞に働いて繊毛の動きを止める。これは一般的神経細胞の興奮と同じで、分子ネットワークに大きな変化はない。ただ、この状況でも光刺激が繰り返すとアセチルコリンが枯渇する。このため新たな合成が始まるまで、幼生の運動は停止する。この結果、幼生はストップ・アンド・ゴーを一定のサイクルで繰り返す。
この時、光シグナルの強さによりもしクロマチン情報が変化し、アセチルコリンの合成速度が完全に変化したとすると、ストップ・アンド・ゴーの感覚を長期的に長くしたり、短くしたりすることができる。即ち、刺激による長期記憶が誕生したことになる。
繰り返すが、これは理論的可能性を述べただけで、実際にどうかはまったく知らないが、神経内の分子ネットワークとクロマチン情報を変化させることで、回路形成なしに神経記憶を形成することができることを理解してもらえたと思う。
この図1,2の記憶システムの基盤の上に新たに回路形成が加わったのが神経記憶で、次回はこの回路形成が加わったことによって生まれる新しい可能性について見ていく。
[ 西川 伸一 ]
神経記憶II
2016年12月1日
前回まで、ゲノム情報を中核とする記憶(Geneme mediated memory:GMと呼ぶ)、クロマチン情報を中核とする記憶(Chromatin mediated memory: CMと呼ぶ)について考えを述べてきた。GMは生命進化全体、CMは生物個体の発生から死までの時間をカバーする記憶システムと言える。
生物学は現象を生物の側から説明しようとして目的論に陥ることが多いが、それを承知で言うと、GMもCMも、過去の経験を記憶し、未来に備えるシステムと言える。
前回見たように、GM、CMそれぞれの記憶システムは階層的に重なって統合されており (前回:図2)、CMはGMを基礎としてその上に形成される。この構造はCMを有する全ての真核細胞で共通で、当然神経細胞も他の細胞と同じように、GM,CMを持っている。このおかげで単一の神経細胞だけでもCMを利用して神経記憶反応を起こせる可能性を前回示した。他の細胞と比べると、神経細胞は刺激に対して早い反応(興奮)を起こすことができるため、神経記憶に見えるが、記憶自体は神経特有というよりCMの機能を反映している。
このGM、CM記憶の上に新たに生まれたのが、神経細胞同士の結合した神経回路形を中心にした記憶(neuron mediated memory:NM)で、これにより動物は、質的、量的に一段高い次元の記憶システムを獲得し、今や地球上に100億個体も存在するという驚くべき繁栄を遂げた人間を進化させる大きな原動力となった。
GM,CMでは、ゲノム情報やクロマチン情報のように分子レベルの媒体が経験の記録を担っているが、NMは細胞間に形成される回路のように細胞レベルの媒体を利用する記憶と言える。神経細胞の特徴は、細胞同士で刺激や抑制の回路を形成できることで、この回路が経験の記録の単位になる。もちろん細胞間のネットワーク形成が見られるのは神経だけではない。例えば、免疫細胞記憶成立には、神経伝達と同じように、サイトカインと呼ばれる分子による、シグナル伝達機構が存在する。また、一つの免疫反応に関わる細胞の種類は、マクロファージや樹状細胞から、何種類かのT細胞、場合によってはB細胞と、神経系と同じように数が多い。しかし、免疫系が神経系と大きく異なる点は、抗原と、それに対する抗原受容体の間の特異的分子結合がネットワークの中心にある点だ。さらに、抗原が常に存在しないと、記憶も維持できないし、反応も起こらない。さらに、抗原刺激とそれを中心にする細胞間相互作用により、それぞれの細胞の増殖や寿命の延長などがおこるため、結果細胞の動態に大きな変化がおこる。
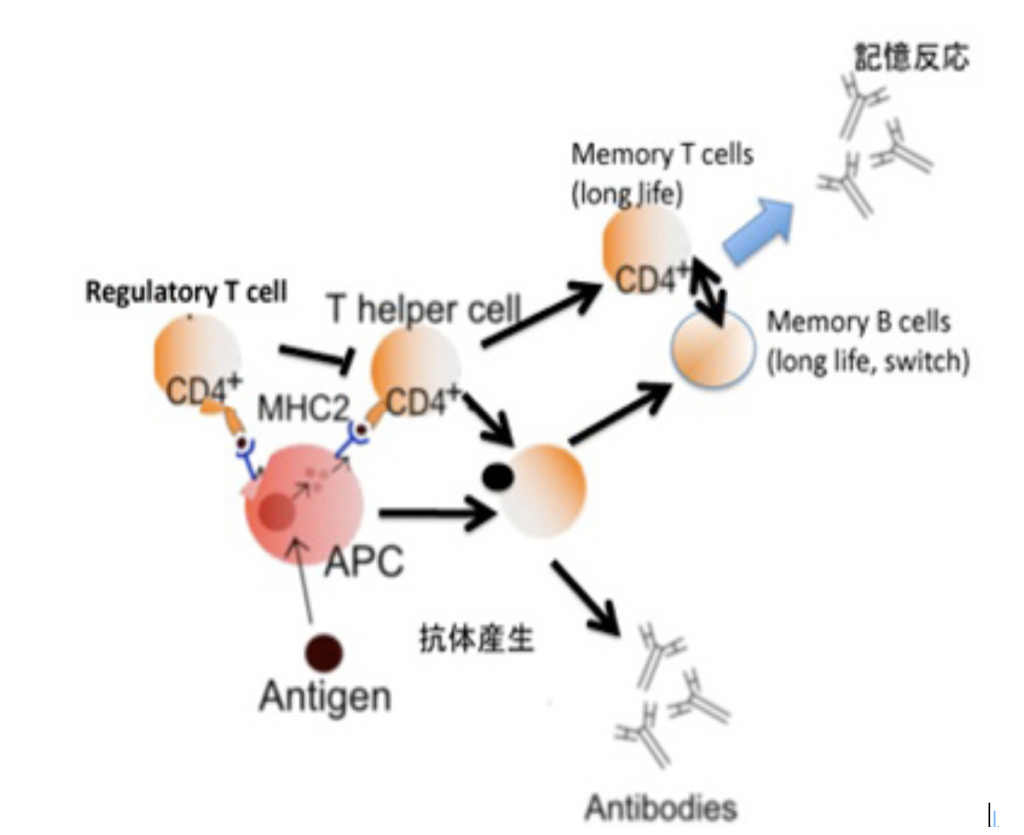
図1 抗体反応での免疫記憶の成立に関わる細胞間相互作用。
図には示せていないが、あらゆる細胞間相互作用に様々な形で抗原とそれを認識する細胞側の受容体が関わっている。
一方、神経細胞回路の場合、匂いや味の感覚のように、外来の化学物質により刺激誘導される場合でも、興奮が神経間で伝達されるときには、最初の興奮を誘導した物質は必要なくなっていることが多い。すなわち、感覚器の興奮が伝えられるとき、感覚を誘導した刺激は必要ない。もちろん刺激の持続時間が短い光などの物理的刺激は、最初から刺激の持続を前提に感覚を伝えることはできない。記憶が維持されるためにも、最初の刺激は必要ない。このように、神経記憶は刺激物質を中心に反応が構成されるのではなく、最初から感覚刺激を複数の神経細胞興奮のパターンへと転換された後、伝達され、記憶される。
どうして最初の刺激を誘導する物質とは全く無関係に、神経回路だけで、記憶が成立、保持できるのかを考えるのが、これからの問題で、詳しく見ていきたいと思う。ただ、神経科学についての私の知識は乏しい。このためこれから書く内容には、間違った記述が含まれる可能性があることを断っておく。
まず、神経細胞回路がどのように記憶に関わることができるか見ていこう。
まず個々の神経細胞をみると、一つの記憶素子として機能できることがわかる。図1に最も単純な電気的記憶素子DRAMを示すが、一個の神経細胞はDRAMに似ていると言える。図に示すようにDRAMは基本的にはスウィッチ役のトランジスタとコンデンサからできており、コンデンサーに電気を貯めることで1 or 0のデジタル記憶が可能だ。
図2 記憶素子DRAMの構造:一般的にトランジスタは電子的スウィッチと考えればいい。
すなわち、スウィッチがオンになって電気がつながれば、コンデンサに電気が溜まった状態になる。一方、スウィッチがオフになると、コンデンサは自然放電して溜まった電気はなくなる。この溜まった電気があるときとないときで、2進法の1か0を記憶できる。
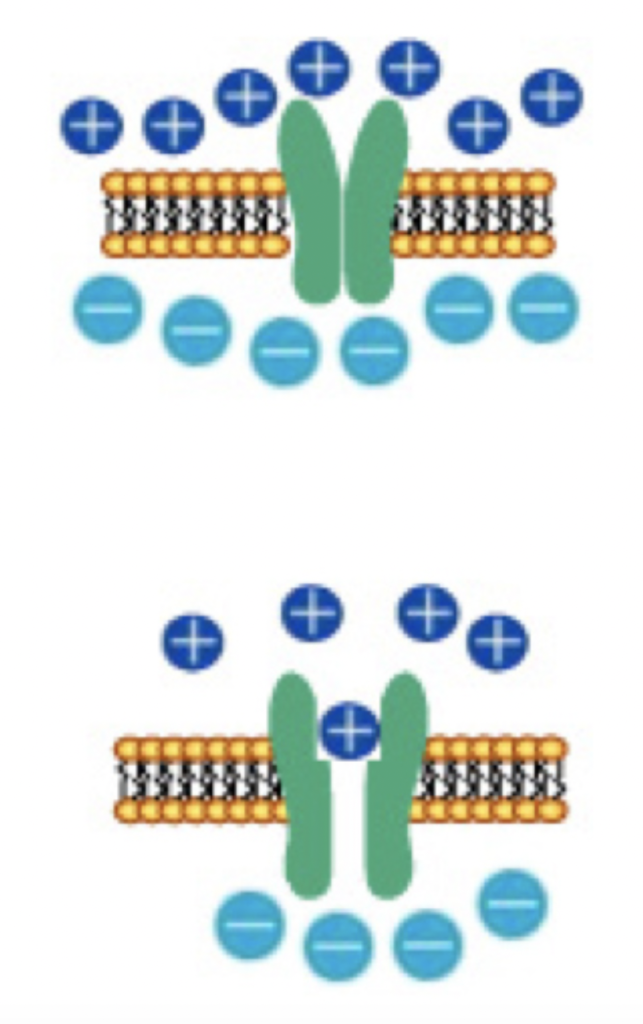
図3:細胞膜に形成された、イオン蓄積システムと、スウィッチ役のイオンチャンネル。
一方、神経細胞はイオンポンプを用いて外界と細胞内の様々なイオン勾配を保つことで、電圧を維持している。細胞膜には特定のイオン分子(例えばNaやK)を選択的に通過させるイオンチャンネルが存在し、刺激によりイオンチャンネルが開くと、特定のイオンだけを勾配に従って流すことができる。すなわち、イオンチャンネルはDRAMでのスウィッチの働きと、イオンチャンネルの特性に応じた一種の抵抗が合わさった素子と考えることができる。
ポンプを使って電位を維持する限り、細胞は自分で発電し、細胞膜という電気を貯めるコンデンサとスウィッチとしてのチャンネルを持つことから、DRAMとよく似た構造をとっているのがわかる。神経細胞の場合、電位はチャンネルが閉じている限り電気維持されるが、チャンネルが開くと外界との勾配はどんどん消えていく。このように原理的には一個の神経が一つの記憶素子として働く可能性を持っているが、実際にDRAMのような使われ方をしているのかどうかについては私にもよくわからない。また、ポンプとチャンネルを備えている細胞は、神経細胞に限らず数多く存在する。したがって、このDRAMに似た構造だけが記憶に大事だとは言えないだろう。
神経細胞自体を記憶素子として使わずとも、神経細胞を組み合わせて回路を形成させ記憶させることも可能だ。エレクトロニクスでは様々な記憶回路がDRAMのような記憶素子の代わりに使われている。もっとも単純なものは図4に示すラッチ回路と呼ばれるもので、インバーター、ANDとORの演算素子を結合させた後、ANDからの出力に応じてフィードバックさせること回路で、0と1を記憶させることができる。
図4 最も単純なRSラッチ回路:説明は省略
電気回路の講義のつもりはないので、ANDやOR といった選択が行われる素子を繋ぎ合わせれば、それ自体で記憶が可能になると考えて貰えばいい。
しかし同じような回路形成を細胞で実現しようとすると、神経のように特殊な形態と機能を持つ細胞が必要になる。いくら興奮性があっても、筋肉細胞ではこのような回路形成は不可能だ。この回路形成が可能であるという点を念頭に神経細胞の特徴を述べたのが、細胞レベルでの脳研究の始祖、ラモンカハールのニューロン4原則だ。
カハールのニューロン4原則
この原理は1890年に書かれたものだが、なぜ神経だけが、演算回路や記憶回路を形成できるのかの原理について見事に説明できている。このことは、図5に示したカハールの網膜神経組織の模式図からよく理解することができる。
(https://en.wikipedia.org/wiki/Santiago_Ram%C3%B3n_y_Cajal)
図5:カハールと網膜神経結合のスケッチ
すなわち、それぞれの神経細胞は、軸索や樹状突起の神経接合部のみで、しかも決まった神経細胞とだけ結合している。そして、感覚細胞からいくつもの神経を経て脳へとシグナルが一方向へ伝わっていることもよく描かれている。まさに、神経細胞は回路形成のために誕生した。
次は神経細胞が形成する記憶回路について、記憶の研究でノーベル賞に輝いたエリック・カンデルの研究を追いながら見てみよう。
[ 西川 伸一 ]
神経記憶III
2016年12月15日
神経科学の歴史について詳しいわけではないが、記憶を始めすべての脳機能が神経細胞(ニューロン)同士の結合した回路により担われていることを最初に示唆したのはスペインの生んだ巨人ラモン・カハールだろう。前回述べたように(前回図5)、カハールはゴルジ染色法を用いて、一個一個の神経細胞を染め出し、脳が様々な神経細胞のネットワークであることを明らかにし、カハールの4原則を提案した。このことから、カハールを神経細胞ドクトリン(ニューロン説)の最初の提唱者と呼んでいいだろう。実際この4原則に適合するネットワークを形成できる細胞は神経細胞以外にない。
カハールのニューロン4原則に加えて、神経回路を考える時に重要なドクトリンをあと二つ挙げておこう。
一つはすでに前回紹介したが、神経細胞は共通のメカニズムで、細胞膜上で起こる電気的興奮を伝搬することができる点だ。静止状態では、神経細胞内外のナトリウムイオン(Na+)勾配によって、神経細胞内外の電位差(静止膜電位)は-90mVに保たれている。外的な刺激でこの電位が変化してある閾値を超えると、イオンチャネル(電位に応じて開閉するNaチャネル)が急速に開き、結果急速にNa+が細胞内に流れ込み、膜電位の極性が変わる(+20mV:脱分極)。この電位の変化は、イオンチャンネルを介してK+を細胞外へ逃すことですぐに元に戻るが、刺激が繰り返されるとイオン勾配が消失して、興奮できなくなる(不応期)。このイオン勾配を復元するためには、それぞれのイオンに特異的なポンプが働く必要がある。このイオンドクトリンに従う細胞であれば、いかに複雑な形態をもち、神経間の興奮伝達に様々な化学物質が使われる場合も、神経を点から点に張り巡らせた電線と同じように単純化して考えることができるようになった。
最後のドクトリンが、神経細胞間の興奮伝達がシナプス接合部での化学物質の伝達によって誘導されるとするシナプスドクトリンだ。すなわち、化学物質の伝達によって、それを受け取る側の神経細胞の興奮を誘導できるが、この場合は電位依存性チャネル(voltage gated channel)ではなく、シナプスで興奮した細胞から分泌される伝達物質と結合することで開くチャネルにより、もう片方の細胞に興奮が伝わることがわかっている。シナプスドクトリンは、神経興奮伝達が一方向にのみ進むというカハールの原理を説明するだけでなく、使われる化学物質を変化させることで、神経間の結合に特異性を与え、また次の神経の興奮に対して促進的にも抑制的にも働けることが説明できる。この二つの性質を合わせると、ほとんど無限の神経回路を作れることが直感的にわかると思う。
この3つのドクトリンを基盤にして、神経回路による記憶維持の細胞学的、生化学的メカニズムについての研究が始まったが、この問題を分子レベルまで掘り下げて説明するのに成功したのがコロンビア大学のエリック・カンデルで、彼はこの研究によりノーベル医学生理学賞に輝いている。
カンデルが記憶回路の解明に用いた動物はアメフラシ(図1)で、大きな殻のないカタツムリと考えて貰えばいいだろう。

図1 アメフラシ(http://en.wiktionary.org/wiki/File:Aplysia_dactylomela_2.jpg)
カンデルは、図2に示した、アメフラシの水管(水を吐き出す管)を刺激すると、エラを引っ込めるという反射に注目した。図2は2000年カンデルがノーベル賞を受賞した時のプレスリリースに掲載されたアメフラシの図だが、腹側にエラと水管が存在している様子がわかる。カンデルたちは、エラを引っ込める反応をアウトプット、水管を触れる刺激、および尻尾への強い電気刺激をインプットとして、それぞれの刺激間の関係や、刺激のパターンが記憶されるかについて調べた。
図2:カンデルの用いた実験系。(C) The Nobel Assembly at
Karolinska Institute
例えば水管を刺激し続けると、アメフラシは刺激に慣れてエラ反射を起こさなくなる(Habituation)。あるいは、水管に触れる前に尻尾に強い刺激を与えると、水管を触れた時のエラの反応が増幅する(Sensitization)。このサイフォン刺激や尻尾への強い刺激が記憶されているかどうかは、一定のインターバルをおいて、同じ刺激を加え、エラが同じ反応を示すかどうか調べることで確認出来る。このインターバルが短い時の記憶が短期記憶で、長い時の記憶が長期記憶になる。カンデルたちは特にSensitization(感作)と呼ばれる行動を用いて短期記憶、長期記憶について研究を行った。
もちろん短期記憶、長期記憶が存在することは、様々な神経システムで観察することができる。しかし、記憶に関わる神経回路を、一個一個の細胞レベルまで分離することは簡単でない。しかしアメフラシの回路では、神経が大きく、また数が少ないため、エラを引っ込める行動とともに、それに関わる単一の神経細胞の興奮を長期間記録し続けることができる。また、刺激後何日もたって、再度同じ神経を探し出して記録することも可能だ。記憶のメカニズムを、行動、神経生理学、そして細胞内での生化学的反応へと還元したいと考えていたカンデルにとっては、アメフラシは選びに選んだモデル動物だった。
図3はカンデルたちが分離した、水管反射の記憶に関わる神経回路の模式図だ。水管への刺激を感じる感覚神経と、エラを引っ込める運動神経がシナプス結合を形成しており、これがメイン回路になる。そこに、水管の刺激をフィードバックしたり、フィードフォワードする刺激性介在ニューロン、抑制性介在ニューロンが結合してサーキットが形成される。このサーキットに、尻尾の感覚神経と結合する調節介在ニューロンが結合して、体の他の部分の感覚を伝えるまさに回路が形成されている。
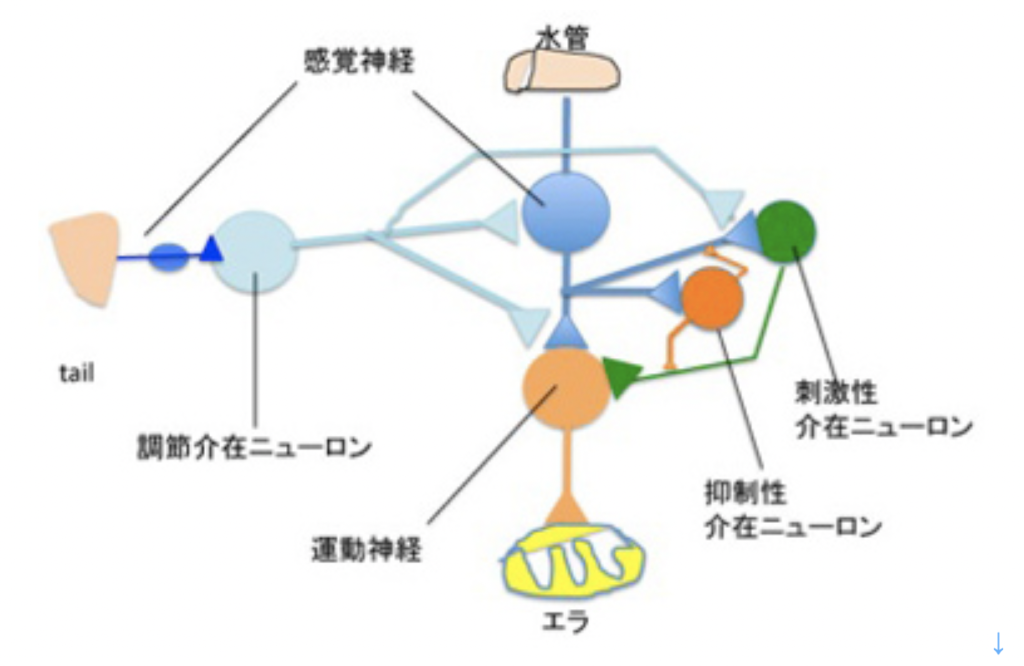
図3 カンデルたちが研究したエラを引っ込める行動を調節する神経回路。実際には、他にも調節介在ニューロンが関わっていることがわかっているが、ここでは省略する。基本的に、水管の刺激を感じる感覚神経と、エラを動かす運動神経の結合した経路に、抑制性および刺激性介在ニューロンが結合し、また全く異なる組織(tail)からの刺激が調節介在ニューロンを介して、この回路に伝えられる。
それぞれの神経は、単一細胞レベルで興奮を記録するとともに、物質を注入することができる。これにより、行動を神経生理学、そして最終的に神経内部での分子間相互作用へと還元することが可能になった。
カンデルたちの期待通り、現在ではSensitizationについての短期記憶、長期記憶のメカニズムはほぼ明らかになったと言える。両記憶のメカニズムについて、同じくノーベル財団のプレスリリースで使われた図を用いて説明しよう。
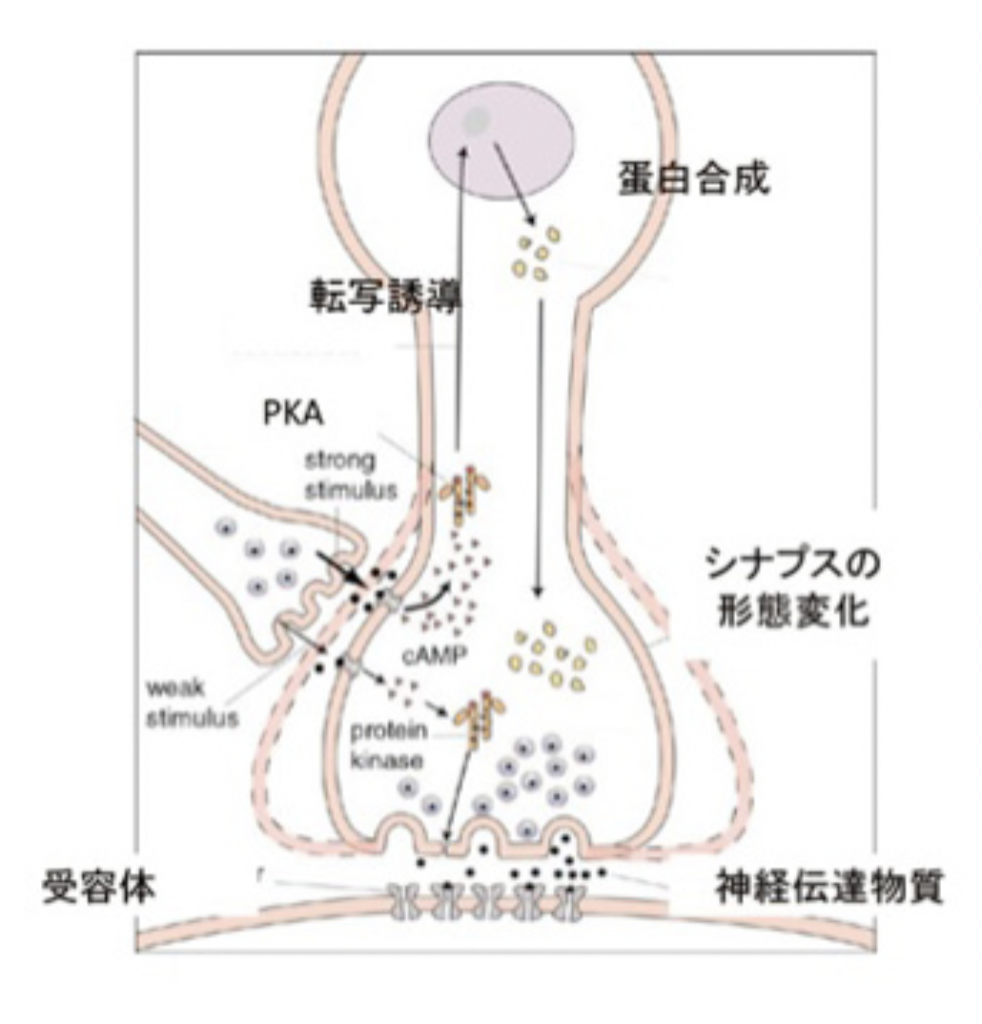
図4:短期記憶と長期記憶。(C) The Nobel Assembly at
Karolinska Institute:説明は本文
結論的には、「短期記憶も、長期記憶も、アウトプットとしてシナプス伝達の強さや閾値を変化させる細胞内の変化」とまとめることができる。
まず短期記憶から見てみよう。尻尾からの調節性介在ニューロンは、水管の刺激を感じる感覚神経と運動神経とを結合しているシナプスに結合している(図3)。この介在ニューロンが興奮すると、1)セロトニンを分泌し、2)このセロトニンがシナプス上の受容体に結合すると、3)アデニルシクラーゼが活性化し、4)cAMPが作られ、5)cAMPにより細胞内のタンパク質リン酸化酵素PKAが活性化され、6)カリウムチャンネルや様々なシグナル分子をリン酸化して、7)より多くの神経伝達物質を分泌するシナプスに変化する、という順序で運動神経への刺激が増強する。ただ、この増強は一過的なPKAリン酸化活性の増強によるもので、時間が経てば元に戻る。
これに対して長期記憶は、同じように活性化されたPKAが一過的にシナプス機能に関わる様々な分子を活性化するだけでなく、図4にあるように転写因子を活性化し、誘導された新しい分子により細胞の分化が誘導されることで、長期にわたるシナプスの大きさ数の変化が起こることで維持されることがわかった。実際には、セロトニン刺激がPKAを活性化するところまでは短期記憶と同じだが、長期記憶の場合はその後CREB1と呼ばれる転写因子が活性化され、新たな遺伝子を誘導することで、細胞自体が分化し、細胞及び組織レベルの構造変化が起こることで長く続く記憶が成立する。言い換えると、細胞をプログラムし直し、違った性質に変化させることが長期記憶のメカニズムだ。この仮説は、感覚神経へCREB1を過剰発現させることで、刺激なしでも長期記憶が成立するという実験による証明された。
実際にはこの単純な回路では、さらに複雑な短期、長期の細胞変化が起こり、シグナルのインプットとアウトプットの関係をさらに複雑にできることがすでに明らかにされているが、その詳細は省いていいだろう。
アメフラシの神経回路を用いた短期、長期記憶については以下のようにまとめることができる。
以上のことから、長期記憶も、短期記憶も、特に特別なメカニズムを使っているわけではなく、多くの細胞と共通の分子を利用していることがわかる。PKAは多くの細胞でcAMP濃度上昇を感知し、短期・長期の細胞変化を誘導するのに働いている。CREB1についても同じだ。ただ神経の場合、刺激により誘導される最終結果が神経細胞同士の神経伝達の強度や閾値の変化になる。またこの変化も、一過性から、様々な長さの時間持続させることが可能だ。
次に、図3からわかるように、神経同士の結合はほぼ無限に複雑化することができる。しかも、神経細胞は形態的にも機能的にも何十種類も存在するため、回路の複雑性はすぐ天文学的になりうる。そして、神経伝達物質や受容体も数十種類存在するため、これによって可能なシナプスを介する興奮伝達の様式も無限に変化させることが可能になる。
しかしネットワークがどんなに複雑でも、背景にある生化学的原理はほぼ共通であることが重要で、どんな複雑な神経系回路でも、刺激が誘導するのは、回路自体の特性のリプログラムだ。
これまで、私の知識を再整理する意味で、神経生物学にあまりに深入りしたが、勉強はこのぐらいにして、次回は神経回路による情報と記憶の特性について考えてみたい。
[ 西川 伸一 ]
記憶と個性
2017年1月5日
ここまで、ゲノム、クロマチン情報、そして神経細胞ネットワークについて、情報と記憶という観点から検討してきた。今回はこれまでのまとめとして、3種類の情報システムを、個性という観点から整理してみようと思っている。
個性というと、「差異、多様性」だけを思い浮かべがちだが、例えばサルと人間の違いを個性とは言わない。個性というとき、あくまでも人間、あるいはサルの個体間で見られる違いについての話だ。したがって、個性という言葉の中には、種としての同一性と個体としての多様性の両方が含まれている。言い換えると、環境に適応するため種としての共通性を維持しようとエントロピーを低下させようとする方向と、ゲノム変異により個体を多様化しようとエントロピーを拡大する方向がぶつかり合うところに個性が成立している(図1)。

図1: ゲノム変異は常に種内のエントロピーを上昇させ、これに対し自然選択はエントロピーを低下させる方向に働く。この結果、種という制限の中で多様性が生まれ、個性が形成される。
おそらくダーウィンほど(ウォレスもだが)、この個性の理解が進化を理解する鍵であることがわかっている人はいなかっただろう。種という制限の中で、次の世代に伝えることが可能な個性が自然に生まれることを確信できないと、自然選択説はでてこない。
人間に個性があるように、あらゆる動物にも個性があることを一旦認めれば、産業革命下の弱肉強食社会を目の当たりにしていたダーウィン時代の都市のインテリにとって弱者が淘汰され強者が残るという考えは当たり前のことだったはずだ。一方、淘汰自体の概念と比べると、あらゆる生物が子孫に伝えることができる個性を自然に発生させるという考えは、背景にあるメカニズムが全くわからないため、受け入れることは簡単でなかったはずだ。もちろん当時の人も、人間を真面目に観察しておれば、それぞれが個性を持っており、誰一人として同じでないことはわかっていたはずだ。しかしダーウィン進化論にとって個性とは、子孫に伝わる性質でなければならなかった。メンデル遺伝学が登場するのは十年後だ。それでも、種としての共通性を基盤として、生殖過程で自然に発生する変異の蓄積が個性を生み、それが子孫に受け継がれると考えたダーウィンの慧眼には瞠目せざるをえない。
次にこの19世紀の議論を、21世紀ゲノム時代の視点で見直してみよう。
今や何百もの種の全ゲノムが解読されている。ゲノムの大きさ、DNA複製のメカニズムを知れば、大腸菌でさえ一回分裂して2個の大腸菌になるとき、両方のゲノムが全く同じである可能性があるなどと考える人はいないはずだ。すなわち、分裂時、あるいは分裂後に個体ごとに異なる様々な変異がゲノムにおこり、ゲノムは多様化し、個性が生まれる。変異は、1代、2代と世代を重ねることにより蓄積し、個体数の増加とともに多様性は増大する。このとき、増殖を損なう変異がおこると個体の維持ができずに自然に淘汰される。逆に、増殖力が高まって他の個体の個性を凌駕することも可能だ。このように、原核生物の場合、生存と複製に必要なゲノム構造が損なわれない限り、種内のゲノム多様性が発生し続け、こうして生まれた個性が環境にフィットすると、今度はこの個性が優勢になり、新しい個性が種として共通の性質になる。
ゲノムのみが経験(自然選択)を記憶できる媒体である原核生物では、ゲノムの多様化はエントロピーの増加を意味する。こうして増加したエントロピーは、自然選択過程により環境にフィットした個性が選択されることで低下する。この過程が異なる環境で起こると、選ばれる個性は異なるため、それぞれの個性へと種全体が収束することで、環境に応じた種が誕生し、種の数は増加する(もちろん環境が同一だと、種は分化できない)。このエントロピーが増減を繰り返す過程の繰り返しが進化で、すべてのゲノムは38億年前の生命誕生まで遡ることができる。このように、ゲノムに蓄えられた過去の記憶はゲノムの個性に等しい。言い換えると、個性とは過去の経験の記憶により形成される多様性と言っていい。(図2)
図2 個体のゲノムは自然に変異を蓄積するようにできている。これにより個性の多様化が起こる=エントロピーが増大する。環境に適応した個体が選択されると多様性は低下し、新しい性質が共通の性質になる。これが様々な環境で起こると、種は多様化する。
原核生物では、ゲノムの記憶がその生物の個性のすべてを決めていると言えるが、多細胞生物が生まれ、生殖細胞と体細胞が完全に分かれると、個体の個性は、ゲノムの個性とは言えなくなる。このことは私たち人間について考えるとよくわかる。
現在では同じ種でも個体間でゲノムが多様化していることがわり、確かにゲノムの個性が子孫に受け継がれ維持されていることは間違いがない。このゲノムの個性から、病気のリスクや、できれば他の様々な性質を予測したいと、個人遺伝子診断サービスが最近提供されている。しかし、個人の全ゲノムを解読したとしても、それですべての個性を予測できると考える人はいない。受精卵が発生を始めてから死ぬまで、我々は様々な経験を通して、ゲノムでは決まらない個性を作り上げていく。これは経験が記憶され、その記憶が個性となるからだが、この記憶はエピジェネティックスとして総称される。ただ、この時の記憶媒体は、クロマチン情報と神経ネットワークの両方(人間では言語も加わるが)からできている。例えば昔人気を博した一卵性双生児姉妹、金さん銀さんの性格や顔があれほど異なるのは、クロマチン情報の記憶と神経ネットワークの記憶の違いが原因だ。
まずクロマチン情報を介する個性について見てみよう。ゲノムの個性と比べた時クロマチン情報を介する個性を研究する難しさは、個体レベルの個性が一つの情報に代表されるのではなく、細胞一個一個の個性の集合として表現されている点だ。
まず発生過程で、何百種類もの細胞が分化するのは細胞ごとにクロマチン情報の違いが生まれるからだ。ただ、発生過程では、決まった細胞を繰り返し安定に分化させることが要求され、クロマチン情報が間違って多様化しないように設計されている。もちろん、この過程でも間違いは起こり、その間違いが細胞の生存や機能を損なわない限り、細胞の記憶として維持される。以前述べたように、クロマチン情報も細胞分裂を超えて維持することができるため、発生の初期に起こった間違いは、多くの子孫細胞に受け継がれる。このような間違いは、時間を経て問題になる場合がある。
有名な例は、1944年の冬、ナチスドイツに封鎖され極度の飢えに苦しんだアムステルダムで胎児期に飢えを経験して生まれた人たちは、高齢になってからインシュリンの分泌が低下し糖尿病になる。これは、胎児発生時に飢えにさらされて起こったクロマチン情報の間違いが、高齢になってから細胞に影響し始めたことを示唆している。同じように、胎児期にお母さんがアルコールを摂取すると、様々な細胞でクロマチン情報の記憶間違いが起こることが知られている。
幸い、クロマチン情報の記憶形成は間違いが少ないように設計されており(増殖因子や分化因子がきめ細かく細胞の方向性を調節する)、それぞれの細胞でおこる記憶間違いはランダムに起こるが細胞の生存から見て許容範囲内で止まっている。またクロマチン情報の変異は細胞ごとの個性になるため、変異が許容範囲を超える細胞は淘汰されるが、個体全体には影響が少ないことが多い。
同じ臓器で同じ細胞が作り続けられるためにはクロマチン情報は必須で、それを維持することは重要だが、発生後も外界のストレスによりクロマチン情報は少しづつ変化する場合が多い。例えば、老化によりクロマチン情報が変化することが知られている。しかし、このような変化は個々の細胞レベルで止まり、各細胞で全くランダムに起こるため、臓器や個体の機能にとって問題にならない。しかし、この個々の細胞にランダムに蓄積した様々なクロマチン情報の変化が集合したものが、クロマチン情報を介する個性として表現される。
以上をまとめると、クロマチン情報の記憶は、各細胞個別に形成される。この記憶は細胞の種類に応じて同一になるよう設計されているが、どうしても小さな記憶違いが生じて、これが細胞の個性を形成する。そして、この細胞の個性が集まって、個体の個性になるが、ゲノムの個性と異なり、個体レベルのクロマチン情報の個性を一つの情報で代表させることはできない。また、クロマチン情報を介する記憶や個性は、各細胞に限局されており、統合されることはない。個体レベルのクロマチン情報の個性は、あくまでも細胞の個性がただ集まっただけの結果だ。
エピジェネティックスな変化の中には、神経ネットワークを介する情報の記憶も当然含まれる。事実、私たち人間の個性は、脳の個性として表現される部分が多い。例えば、生後訓練を繰り返して筋肉が発達し、早く走れるようになる過程にアメフラシで見たのと同じ神経ネットワークを介する記憶が大きく寄与している。もちろん、筋肉自体も発達し、これにはクロマチン情報の変化も寄与するが、神経系の記憶が存在しないと筋肉は発達しない。残念ながら、こうして訓練した結果は、次の世代に受け継がれず、一代限りだ。
神経ネットワークのメカニズムを突き詰めると、細胞が発生し、成長し、老化する過程で起こるクロマチン情報の記憶と同じと言っていい。どんなに複雑な脳神経系でも、進化過程でゲノムに蓄積された記憶情報に基づいて空間的・時間的に発現する特異的なシグナルに従って順々にクロマチン情報が変化した細胞を分化させ、これをゲノムの記憶に従って立体的に構造化させることで、形成される。また、生前、あるいは生後それぞれの神経が受ける刺激に対しては、一次的な反応が細胞内シグナル伝達経路の活動として反応する。そしてシグナルの質や強さに応じて、前回見たように、神経細胞のクロマチン情報が変化し、長期記憶が成立する。しかし図3に示すように、神経細胞の反応は他の体細胞の反応と全く変わるところはない(図3)。
図3:神経細胞と一般体細胞の比較。
説明は文中。
一方、一般の体細胞と神経細胞の記憶は、個々の細胞内で起こる変化が個体レベルで統合されるかどうかという一点で大きく異なっている。すでに述べたように、クロマチン情報の記憶に起因する細胞の個性は、ただ集合して個体の個性に寄与するだけだが、神経系では細胞レベルのクロマチン情報の変化に起因する個性は、細胞同士がネットワークを形成することで、個体レベルで統合される。この統合は、神経細胞にとどまらず、筋肉やホルモン産生細胞など、多くの体細胞をも統合できる。
実際には、同じような細胞の変化を統合して個体の個性を変化させることは、内分泌システムにも見ることができる。オタマジャクシがカエルへと変態を遂げる時、甲状腺ホルモンが分泌され、体全体の細胞を統合する。同じことは、私たちが思春期に急速に様々な性的特徴を発生させるのも同じだ。しかし、一見個体全体を統合している高等動物の内分泌システムを詳しく見てみると、ほとんどがどこかで神経系とつながっており、神経ネットワークの記憶の支配を受けていることに気づく。
このように、神経細胞は、神経細胞にとどまらず様々な細胞と直接、間接にネットワークを形成する能力を開発し、全く新しいレベルの個体性の統合を可能にしている。
これに加えて、神経ネットワークでは、視覚、聴覚、嗅覚など、外界の変化を迅速に、立体的に捉え、この入力を個体として統合された神経ネットワークにリンクさせることができる。これにより、環境を全く異なるレベルで自己化することが可能になった(例えば景色の記憶は、環境を自己化したと言える)。もちろん、外界の自己化は原始ゲノム発生以来、生物進化の駆動力と働いてきたことだが、神経系による自己化は、あるがままの外界を表象し自己の延長として自己化する点で、これまでの自己化とは全く異なる。
この個体内外の統合と、自己の発生についてはわかりにくいと思うので、次回は、「統合と自己」というタイトルで、神経系ネットワークが可能にする個体と細胞、個体と外界の統合能力について解説してみたいと思っている。
[ 西川 伸一 ]
統合と自己:細胞の個性と個体の個性
2017年1月16日
今回は個体と細胞の関係を考えながら、神経情報の特殊性について見てみる。
教科書的には、多細胞体制の誕生は細胞の個性と個体の個性の分離をうながし、またその結果個体のゲノムを継承するために生殖細胞が誕生したことになる。しかし、生殖細胞が誕生した後も、有性生殖だけが個体のゲノムを継承する方法ではない。例えばプラナリアのように個体全体が分裂する場合もある。植物では体細胞からカルスが形成され、そこから完全な植物個体を形成することができる。

図1:プラナリアは成長すると個体自体が分裂することが古くから知られている。(出典 https://www.flickr.com/photos/internetarchivebookimages/20705306246/sizes/o/)
有性生殖と無性生殖が共存できるのは、ほぼ同じゲノム情報が全ての細胞に存在しているからに他ならない。もちろん、細胞ごとにゲノム情報の小さな違いは生じているが、原則的にほぼ同じゲノム情報が個体の全ての細胞に存在していると考えていい。無性生殖を行わない動物でも、体細胞の核移植によるクローン動物作成実験から(哺乳動物のように複雑な動物でも、体細胞のゲノムを未受精卵に移植すると、新たな個体が発生する)、全ての細胞に完全なゲノム情報が存在していることが示された。この意味で、ゲノム情報の同一性が、細胞と個体の統一性を保証していると言える。
しかし、植物やプラナリアでは体細胞から個体を形成できるのに、私たち人間の細胞は、決して個体を形成することはない。この理由の一つは、私たち人間の細胞は、ゲノムは受精卵と同じでも、体細胞への分化過程で変化した細胞それぞれのクロマチン情報を簡単にはプログラムし直すことが難しいことだ。実際プラナリアも同じで、分化した細胞は他の細胞に分化し直すわけではなく、全細胞のうち3割がクロマチン情報を変化させやすい多能性幹細胞でできているからだ。同じゲノムを持っていても、不可逆的にそれぞれのクロマチン情報が選び取られて発生が進むことを、英国の発生学者ウォディントン(C.H. Waddington)はエピジェネティック・ランドスケープ(epigenetic landscape、後成的風景)という絵を用いて表現している。しかし、分化細胞を他の種類の細胞へと変化させることが難しいお陰で、私たちは明日皮膚が血液に変わってしまっているという心配をしなくて済む。
図1 ウォディントンの描かせたepigenetic landscapeの絵をわかりやすく書き直した図で、この概念を示す際、最もよく使われる。上部にあるビーズは様々な場所に動き得るが、一旦分化の方向性が決まると、他の道は選べないことが示されている。
結局、我々人間では、受精卵から始まる一生で一回きりの、ゲノムの指示に従う発生過程を通す以外に、必要な細胞を準備することはできない。ES細胞やiPS細胞の研究から、受精卵に限らず、体のあらゆる細胞を作る能力を持つ細胞が得られることが明らかになり、他の細胞から区別して多能性幹細胞と呼ぶようになっている。しかしあらゆる細胞に分化できる多能性幹細胞の塊から始めても、正常な個体を発生させることはできない。なぜなら、動物の発生過程では、個々の細胞が個体との関係を読み取り、これにより得られる情報の指示に従ってクロマチン情報を変化させることが要求される。例えば四肢はほとんど同じようなメカニズムで体幹から飛び出すが、頭に近いところにある細胞から腕が、遠いところにある細胞から足ができる。これも、各細胞がボディープランから見て最もふさわしい組織の形成を指示する、すなわち適切なクロマチン情報を確立するための情報が体の各部に用意されているからだ。しかし、体の体制を構成しているのも細胞だ。どちらが原因で結果かがわからないという因果性の問題が発生学には常に存在している。
ゲノムには、発生に応じて必要な遺伝子セットを発現させるためのプログラムが存在し、多能性の幹細胞は発生に合わせて様々なクロマチン情報記憶を展開することができる。多能性幹細胞は様々な条件下でほとんどの細胞へと分化できるが、秩序ある個体を形成するためには細胞の多能性だけでは不十分で、細胞が初期胚が持つ個体の体制からのシグナルを受けることが必要になる。例えばES細胞を集めて個体に移植したり、あるいは試験管で培養したりしても、奇形腫と呼ばれる秩序のない構造ができるだけで、マウスはできない。しかし同じES細胞を胚盤胞に注入すると、胚盤胞の体制に組み込まれてES細胞は正常個体を形成することができる。
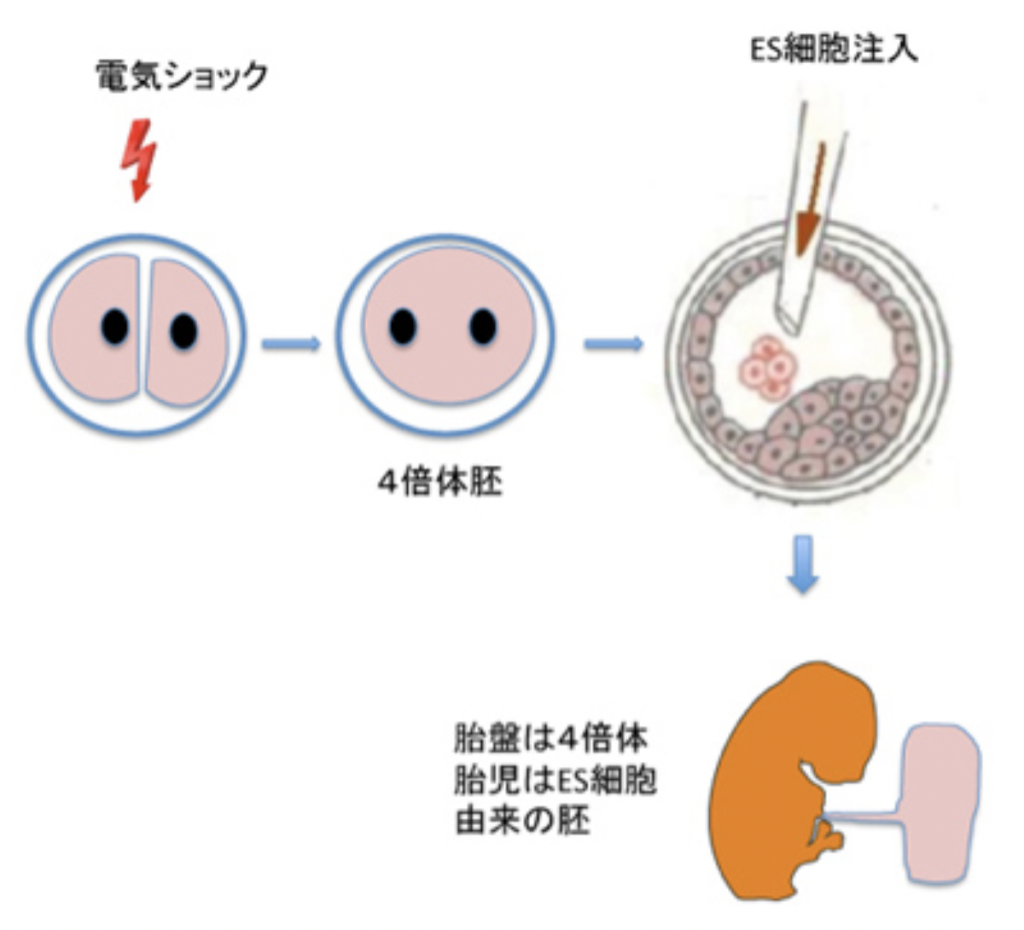
図3 4倍体補完法の説明 説明はテキスト参考
胚盤胞の体制自体が持つトップダウンの力を最もよく示す例が4倍体補完法と呼ばれる方法だ(図3)。受精卵が分裂した時に電気ショックで細胞を融合させると4倍体の胚ができるが、この胚盤胞をES細胞の塊に指示を与える構造として用いることができる。4倍体の胚は胚盤胞期まで発生するが、胎児細胞はそれ以上発生ができない。一方胚外のトロフォブラストは機能的胎盤を形成できる。このため胚盤胞に移入したES細胞やiPS細胞は、死滅する4倍体の細胞の代わりに発生する。すなわち、胎盤は4倍体の細胞から、胎児部分は注入したES細胞からできた合成胚を作り、発生させることができる。単純な構造に見える胚盤胞にも、細胞の分化に指示する体制が存在することがわかると思う。
このように発生学は、細胞の分化を指示する個体の体制とは何か?そのシグナルを受けた分化細胞がどのようにして個体へと統合されるのか?など、常に全体と部分の関係を研究してきた。このことは、例えば古典的な発生学で、オーガナイザー(organizer)とか、誘導物質(inducer)、ボディープラン(body plan)という個体の体制を示す言葉が重視されていることをみれば理解できる。この発生学の特殊用語を使ってまとめると、ゲノムの記憶を正確に展開するためにも、最初はオーガナイザーが必要で、このオーガナイザーもまた細胞からできており、ゲノムに指示により形成されるのだが、卵の形成過程、受精過程、着床過程など外部の要因が関わって形成される。
このように「卵が先か、鶏が先か?」の問題と同じで、発生では全体と部分は一体化しており、原因と結果の区別を困難にする。同じように、発生でのゲノム情報とクロマチン情報の関係も一体化している。発生でのクロマチン情報の変化は、ゲノムにコードされたプログラムで進むが、クロマチン情報がないと複雑な多細胞生物のゲノムの記憶を展開することができない。
では前回述べた多細胞動物に見られるゲノム情報、クロマチン情報の個性とはなんだろう。発生過程で一つの系列内の細胞の多様性が生まれることは、ゲノムやクロマチン情報の間違いと考えていい。これらの情報の変異は細胞ごとに起こるが、許容範囲以上の間違いは致命的になるか、細胞レベルで除去される(例外と言える免疫系は後で説明する)。発生、成長の過程で個々の細胞レベルで発生する個性は、個体の個性として統合されることはないが、細胞レベルの個性が個体に影響を及ぼす時、病気、老化などの異常として現れる。
少し長く説明しすぎたが、神経情報が誕生するまでは、新しく生まれた情報システムも、それ以前に存在した情報から独立することができていなかったことを示すためだ。これに対し、新しく生まれた神経情報は、他の情報に依存した生命活動から、大幅な独立を勝ち取っている。
もちろん神経ネットワークも、その形成や維持はゲノム情報、クロマチン情報に完全に依存しており、神経情報だけが独立することはない。体がないと、脳は機能せず、神経情報維持できず、心もできない。しかし、神経情報のオペレーションを詳しく見てみると、それ自体で他の情報から独立して働くことができ、また各細胞に生じた個性がそのまま神経系全体の個性として統合されていることがわかる。
例えば、一卵性双生児間の個性を考えてみると、細胞レベルのクロマチン情報の違いが集まって生まれる個性の違いと比べ、神経ネットワーク情報に起因する性格や記憶の違いは際立って大きい。すなわち、神経ネットワークの枠組みが一旦形成されると、神経情報のオペレーションは自由度が高く、結果多様な個性を形成できることを意味している。個性が多様化できるということは、他の情報からの独立性が高いことだ。この独立性が、デカルト以来私たちが心身二元論に陥る理由の一つだと思う。
しかしなぜ身体の死とともに消滅する運命の神経ネットワークの情報が、心身2元論という錯覚を生むほどの独立性を獲得できたのか?その原因について最後に考えてみよう。(以下は私の妄想でしかないことを断っておく。)
神経ネットワークが持つ最も重要な特徴は、様々な神経細胞があるにもかかわらず、各神経細胞の興奮、細胞内(軸索や樹状突起)での興奮の伝搬、そして神経間の興奮伝達のための原理がほぼすべての神経細胞で共有されている点だ。そのおかげで、視覚であれ、嗅覚であれ、触覚であれ、全てのインプットを同じ神経ネットワークで共有することができる。また、インプットに対するアウトプットも、筋肉を動かす運動からホルモンの分泌に至るまで実に多様で、多くの組織を直接間接に支配してネットワークの中に組み込むことができる。もちろん、インプット・アウトプットだけでなく、神経同士が刺激・抑制し合う回路も形成される。同じ原理で作動できるため、神経細胞の数さえ増やせば、無限に神経細胞同士を組み合わせ複雑なネットワークを形成することができる。こうしてできた神経ネットワークも、神経間の結合の特異性や強さを変化させるためにはゲノムやクロマチン情報に依存したメカニズムを用いるが、クロマチン情報が「ゲノムのon/off」のための情報であるのとは全く異なり、形成される記憶自体は、直接ゲノムやクロマチン情報と関係はない。
このようにゲノムとクロマチン情報に依存して発生が進み一旦回路が形成されると、あとは外来の刺激が回路のパターンを変化させる例は、すでにアメフラシの水管反応で説明した。アメフラシ水管反応回路では、刺激を受けなかった回路と、刺激を受けた回路では全く回路特性が異なる。これは回路構造の中の神経細胞の特性が変化したためだが、この変化は回路全体の個性として統合される。このように、神経回路はこれまでの情報と比べて、高い自立性を持ち、興奮伝達の原理を共有することで無限に複雑化する可能性を獲得した。しかも、さらにこの共通性のおかげで、個々の細胞レベルで生じた神経細胞の個性はネットワークにより共有されることで、神経系全体の個性へと統合される。
細胞の個性を統合することができる神経系の誕生は、刺激が消えた後もその刺激の影響を維持することを可能にした。この点については、同じようにほぼ無限と言える外来抗原に反応できる免疫系と比べてみるとよくわかる。
免疫系は無限の化合物に対して反応できる。これは、多様な抗原の一つ一つに対して反応する個別のリンパ球が体内に存在するからだ。すなわち、同じ細胞を繰り返し作り、細胞に個性が生まれるのを極力拝する他の組織と異なり、リンパ球は最初から異なる個性(抗原特異性)を持つよう分化する。この個性はゲノムレベルの個性で、抗体やT細胞受容体遺伝子を不可逆的に変化させ、それぞれのリンパ球に異なる抗原受容体遺伝子を発現させるリンパ球特異的な遺伝子再構成メカニズムに依存している。しかし個々の細胞レベルの免疫反応は抗原に反応する細胞だけでとどまる。すなわちリンパ球同士の反応には必ず抗原の介在が必要で、抗原がなくなると細胞間の相互作用は消滅する、一方、神経の場合は刺激が消えても、細胞相互作用は維持できる。
ニールス・イェルネらは、抗体が他の抗体により認識できる(イディオタイプ)という現象を元に、リンパ球同士は抗原を介さずに、神経のように直接相互作用できるとするイディオタイプネットワーク説を唱えた(図4)。しかし、抗体が認識し合ってネットワークを作るという考えはは最近ではあまり支持されなくなっている。結局、免疫系が無限の抗原に反応できるのは、ネットワークとは無関係に、リンパ球の抗原反応性に最初から多様性があるからだ。結局免疫反応とは、抗原反応性の異なる個別の細胞ごとに増殖、分化寿命延長などの細胞学的変化が起こることに他ならない。
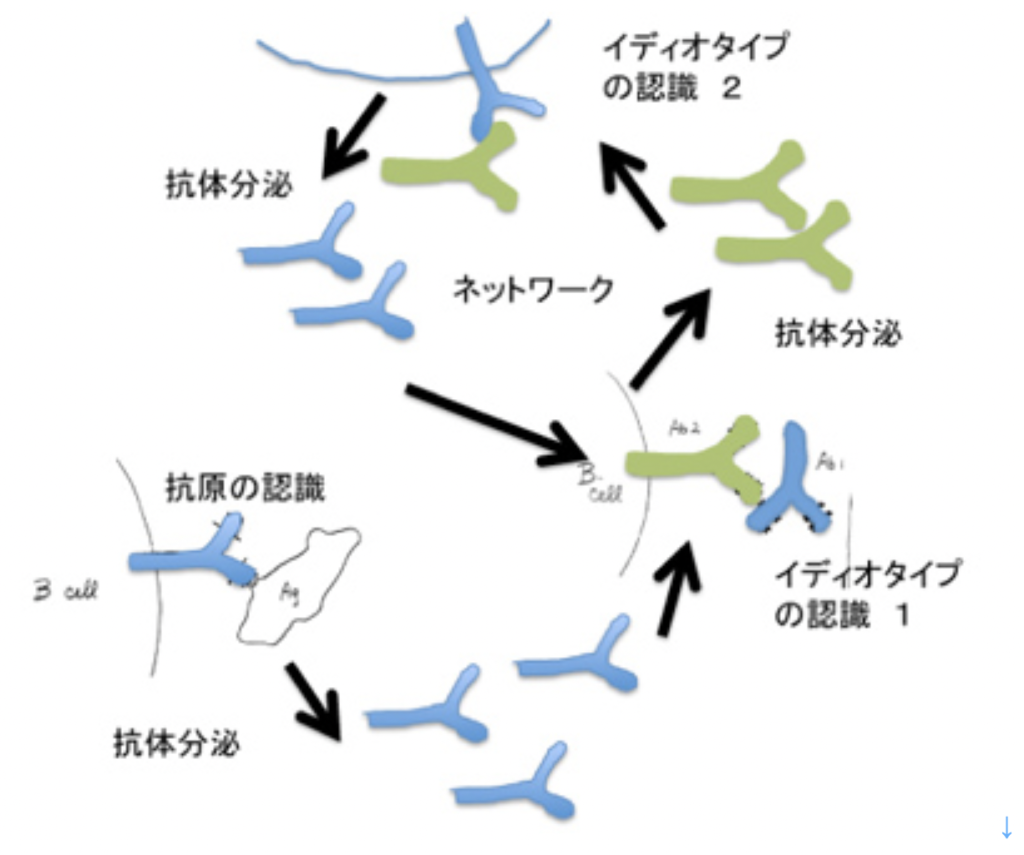
図4 ニールス・イェルネらが考えたイディオタイプネットワーク説。彼のノーベル賞受賞論文を元に書き直した。最初抗原により刺激されたB細胞から分泌された抗体1(空色)の抗原結合部は(他の抗体分子とアミノ酸配列が異なるため)抗原としても働き、抗体2(緑色)により認識される。この相互刺激関係が成立すると、抗原なしに刺激は持続する。
一方、イェルネが考えたように、免疫系でネットワークが成立するためには、最初の刺激が消えた後も、リンパ球同士で刺激しあえるメカニズムが必要だ。免疫系では、図4に示したことは理論的には可能でも、現実には特殊な状況だけで起こっているようだ。
一方図4に示したネットワークは神経系では成立できる。神経系も、味覚や嗅覚として化学物質を感じ、記憶できる。化学物質に反応する点では、免疫系と同じだが、例えばワインを一口飲んで、実際の刺激は胃の中に消えてしまっても、味や匂いを他の感覚と関連させ、記憶として維持することができる。これは、刺激が消えた後も、匂いや味の刺激によって生まれた新しい回路の活性が維持できることを意味している。これが可能なのも、神経細胞が同じ原理を共有し、回路を形成できるからだ。
以上、全ての神経細胞が興奮のための原理を共有することで、神経細胞同士で無限に複雑なネットワークを作り、また各神経細胞の変化を、ネットワーク全体の変化として共有できるようなったことで、心が身体から離れてしまうという心身2元論の幻想が生まれるほど、神経ネットワークによる情報が、ゲノムやクロマチン情報から独立した、独自の情報系を作ることができたことを納得していただいただろうか。
次回からは、各細胞の変化が統合可能で、他の情報から独立している神経情報系により可能になった脳新しい情報機能について見ていきたい。
[ 西川 伸一 ]
ゲノムの発生学 I
2015年10月15日
ゲノムの解剖学、そしてゲノムの働き(生理学)とくると、当然次ぎにくるのはゲノムの発生学だ。ゲノムの発生学のほとんどは進化の過程とオーバーラップするはずで、この紙面で簡単に説明することなど叶うはずがない。代わりに今回からゲノムの発生学として、これまであまり議論されてこなかった観点からゲノムの誕生や進化の過程をゆっくり整理し直したいと思っている。今回は、そのイントロダクションになる。
重要なことは、ダーウィン時代と異なり、ゲノムの解剖学や働きについてずいぶんわかっている点だ。従って、ゲノムについて理解が進んだ時代のゲノム発生学(進化学)とは何かを考えることになる。これまでゲノムの解剖学や生理学の説明を通して、ゲノムは多様な内容を表現していても、結局は情報であるとことを強調してきた。この意味で、ゲノムの発生学を考えようとすると、情報の発生学を考えることになる。ところが日常情報という言葉を当たり前のように使っている我々も、情報の発生学と言われる何を考えていいのか戸惑う。なぜだろう?まず生物のゲノムを私たちが普通使っている情報との比較をしながら考えてみよう。
生物が生きて活動するオペレーションのためにゲノムが表現している情報には、アミノ酸配列の情報、機能的RNAの配列情報、遺伝子発現を調節するための様々な情報、遺伝子の円滑な発現に必要なゲノム構造化のための情報など、多様だ。ただゲノムを分解してみると、ゲノム上の情報のほとんどは、全て生命オペレーションに必要なタンパク質と様々な機能的RNAを必要に応じて作るために組織化されている情報と言える。このため、生物を機械にたとえ、ゲノムを生物の設計図と説明する人がいる。例えばゲノムについてかかれた本を見てみると、「初めて学ぶゲノム生物学:生命の設計図」、「ゲノム:命の設計図」、「ゲノム=人間の設計図をよむ」などのタイトルが踊っている。個人的には設計図より、パソコンのソフトウェアのように、生物のオペレーションの指令書といった方がまだ正確なように思えるが、なぜ私たちはゲノムを設計図や手順書に似ていると考えるのだろう?
デカルトは人間を心と体に分け、体は機械と同じだと考えた。また心のない動物は機械でしかないとも考えた。そしてこのドグマはほとんどの生命科学分野の根幹を今でも脈々と流れている。しかし、実際の生物は機械とは全く違っていると誰もが直感する。何が違うのか?
自分で考えて将棋を指すように見えるコンピュータといえども、機械はその背景に設計者、すなわち人間の意図が存在している。例えばシャベルカーを考えよう。シャベルカーは穴を掘るという人間が決めた目的に従って設計され、組み立てられる。そして完成したシャベルカーを運転してその意図を実現するのも人間だ。動いていない機械だけを見ているとそこに人間の存在はないが、実際には機械そのものが人間の意図の塊だ。石より固い鉄、自由な動きを可能にする油圧システム、駆動エンジンなど、本来はシャベルカーとは全く別に開発されてきた数多くの部品が、シャベルカーに集められている。全ては設計者(=人間)の意図に従って組み立てられるが、シャベルカー位複雑な構造になると、この意図を情報として表現する指示書や、設計図が必要になる。さらにこの機械を設計するのも人間であることから、運転する人間が扱いやすいよう設計するのも重要だ。
図1 あらゆる機械はその背景に人間の意図や目的が存在し、それに合わせて設計される。
現代なら、人工知能を導入した自動のシャベルカーも存在しているはずだ。自分で学習し、初めての場所でも上手に穴を掘ることができるかもしれない。しかし、一旦人間により決められたシャベルカーの目的は変わることはない。シャベルカーが自分で目的を逸脱してスポーツカーになることは決してない。これは、人間が操縦しようと、全自動であろうと、全ての機械の目的は人間により決められており、またそのオペレーションも人間との対話の上で行われるよう設計されている。
少し余談になるが、最近自ら考え判断する人工知能の開発が進められているが、これが成功するといつか人間から独立し、人間を支配するのではないかと心配されている。ただ人工知能も機械である以上最初の目的は人間が与える。このとき心配する必要があるのは、与えられた目的をもっとも有効に成し遂げようとして、人間を排除することだ。例えば人工知能に穴を掘れと命じて、その後途中で穴掘りをやめさすためスウィッチを切ることを考えてみよう。もし人工知能がこのやめるという意思を、目的遂行を邪魔すると判断すれば当然スウィッチを切ろうとした人間は排除されるだろう。とはいえ、機械はその背景にある人間の決めた意図なしでは存在しない。
この機械の背景にある人間の意図と同じものを、私たちは生物にも感じる。この感覚が、私たちにゲノムは生命の設計図と言わせている。もしデカルトに、生物という機械に外から目的(設計図)を与えているのは何か?と直接質問したら、おそらく彼は生物に目的を与え、それに沿って設計するのは神だと答えるように思う。すなわち、機械の背景に必ず人間がいるように、生物という機械には神の意志が存在すると考えていた。しかし、神を生物の背景から排除すると(私は当然だと思っている)、機械に対する人間のように、生物に目的やデザインを与えるものはどこを探しても見当たらなくなる。実際にはこの生物の目的の探求が、18世紀自然史思想の誕生以来続けられ、ダーウィンを経た後ようやく見えてきた解の一つがゲノムに集約した。この歴史を考えると、生命の構成成分をコードする遺伝子が組み込まれたゲノムを設計図や指令書と考えて不都合はない。DNAという媒体は物質だが、ゲノム自体は情報で、物質的ではないし、目にも見えない点でも、機械の背景にある人間の意図に似ていると言えるのではないだろうか。
しかし、ゲノムを生物の設計図や指令書であることを認めたとしても、機械の設計図とは全く違う問題がゲノムにはある。それは書いた人がいないという問題だ。私たちは普通の情報を考える時、それがどうしてできたか、すなわち発生学はあまり問わない。というのも、情報には確実にそれを発信した人間がいるからだ。機械のデザインや設計図も同じように、書いた人がはっきりしている情報だ。一方設計図と言っても、ゲノムには書き手がいない。このため、ゲノム情報を考える時は、書き手がいないのに情報がどうして生まれるのかをまず知る必要がある。
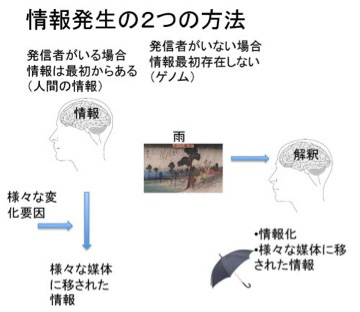
図2 普通情報にはその発信者がいる。しかし、発信者のいない現象も、解釈という過程を介して情報化することができる。
私たちが情報について語るとき、必ずしも発信者のはっきりした情報だけが念頭にあるわけではない。例えば、私がこの原稿を書くのに疲れて窓を見ると、天気は秋晴れだ。ちょっと散歩でもするかと伸びをして部屋から出たとしよう。これは、窓の外の日光の様子を私が心地よい秋晴れと解釈した結果だ。逆もある。例えば雨が降っていたら、傘を持って外に出る。これも雨が降っているという情報を私が傘を持って出ることを指示する情報と受け取ったからだ。この場合私が情報として受け取った天気の状態は自然現象で情報ではない。同じ雨でも、畑にとって恵みの雨もあれば、川の氾濫を引き起こす雨もある。全て状況と解釈者の状態によって決まる一種の解釈だ。すなわち、誰かが書いた情報でなくとも、それを解釈する主体があれば情報になる。発信者のいないゲノムが情報になりうるのは、この解釈という能力がゲノムに備わっており、この過程を通してゲノムが情報化していくと考えられる。
この解釈プロセスは、原始生命誕生時のDNA分子が情報を担う様になりゲノムが誕生する過程と、ダーウィン進化過程に分かれる。次回からできるかどうか自信はないが、この過程についてさらに考えてみたい。
[ 西川 伸一 ]
ゲノムの発生学II
2015年11月2日
物理化学現象を人間が解釈するとき現象の情報化が起こるが、本来物理・化学に情報という概念はない(量子力学は少し忘れよう)。従って、生命が誕生するまで、物理化学法則のみが支配する地球に情報は存在しなかった。最初の生命については知る由もないが、生命誕生とほとんど同じ時期に(同時という意味ではない)、DNAを媒体とするゲノム情報が生まれたことは間違いない。これが地球における最初の情報の誕生になる。ゲノムの発生学はもちろんこの誕生から始まる。自信がないが、地球上での情報の誕生について今回は考えてみよう。
発信者なしに情報が生まれるためには、もともと情報でないものを情報として解釈する「主体」が必要になる。ただそんな主体は生物誕生前にはもちろん存在しない。最初に存在したのは物理化学法則に従う分子と分子の相互作用だけだ。しかし、分子間相互作用も一つの分子の側から見てみると、近づいてきた分子と結合するか離れるか、自分の構造に基づいて解釈していると言える。したがって、解釈という行為自体は核酸のような情報分子に限らず、どの分子にも考えられる。しかし私たちの住む大気の中で、酸素、炭酸ガス、窒素は常に衝突しているが、それぞれが反応する(解釈する)ことはほとんどない。すなわち、解釈のような振る舞いは、複雑化した分子構造のほうが生まれやすい。特に有機分子のポリマーは様々な分子と相互作用が可能だ。タンパク質や核酸が様々な高分子と「非特異的」に結合することは、生化学実験の経験者は誰もが知るところだ。
次に考える必要があるのは、この条件を満たす有機ポリマーが原始の地球に存在していたかどうかだ。生命に頼らず有機物を合成する有機化学はヴェーラーの尿素合成までさかのぼる。ただ、生命誕生に関わる化学の先駆は、ユーレイとミラーのアミノ酸の合成実験で(図1)、この研究以後生命誕生の化学について地道な研究が進んできた。
図1 Urey-Millerのアミノ酸合成実験。水蒸気とメタン、アンモニア、水素を混合した反応容器に、雷を模した放電を行い各成分の反応を誘導、得られた化合物を水にトラップし、また加熱して反応容器に循環させる。この方法で様々なアミノ酸が合成できることを示した歴史的実験(Wikiコモンズより)。(Science 130, 245, 1959)
この実験以降に蓄積されたデータを詳細に解説するのは私には荷が重い。都合のいいことに、これまでの研究をまとめると、核酸、アミノ酸、脂質など有機化合物は生命以前の地球で合成されていた可能性が高い(合成できていないと生命は誕生できないことから当然だが)。また、自由エネルギーから見たとき、簡単には起こりえない有機分子のポリマー形成も、熱、濃縮、氷結などの変化が、鉱物や粘土などの自然の触媒と組み合わさると可能であることが示されている。例えば、モンモリロン石は、アミノ酸の重合と、リボ核酸の重合を促進する触媒活性を持っており、アミノ酸、核酸のポリマー形成を誘導できることが示されている。従って、アミノ酸や塩基が重合した様々な長さのポリマーが原始の地球に存在しできた(このabiogenesis, systemic chemistryについては優れた本や総説が書かれており、とりあえずKepa Ruiz-MirazoらのPrebiotic systems chem.istry: new perspectives for the origin of life, Chemical Review 114, 285-366, 2014 を紹介しておく。)。
次の問題は、様々な有機分子が存在するようになった地球で、情報、特に核酸を媒体とする情報が生まれた過程だが、これについては全くわかっていないし、実際に起こったことを特定することは永遠にできないかもしれない。しかし様々な可能性を考えることは可能だ。一つの可能性を考えてみるが、すべて私の頭の中妄想だと思ってほしい。
明確な意図やデザインの元に合成されたわけでなくとも、アミノ酸や核酸がランダムに繋がって生まれた分子同士が、互いに相互作用できることが知られている。しかし相互作用と言っても全ての相互作用は偶然の産物で、ただ分子同士が付いたり離れたりしているだけだ。ただランダムにでも多様性が生まれると、その中から一部が選択され進化が起こることを思い出してほしい。環境による選択は多様な分子反応の間でも起こり、その結果反応が向かうべき目的のようなものが生まれる。すなわち、ダーウィンの進化論を生命のない分子にも適用することができる。例えば分子の安定性と持続性という目的が自然に反応している分子の間に生まれることは十分考えられる。他の分子より高い安定性があると当然存在時間が長くなる。分子の存在時間は環境による選択の結果として現れ、ダーウィン進化での「長い首」と同じ目的性が生まれる。
このような選択が起こることを一番理解できるのがホモキラリティーという現象だ。例えば地球上の生物に存在するタンパク質はL-型アミノ酸だけからできている。一方糖鎖になるとD-型だ。ランダムに合成されるアミノ酸の中からなぜ生物はL型のみを使うようになったのか?すべての生命でこの選択が行われていることから、おそらく生命誕生以前のabioticな過程で、ホモキラリティーが生まれたと考えられ、そのメカニズムの研究が進んでいる。メカニズムはともかく、この現象から生命の関与なしにこのような選択が生物界全体で起り得ることを理解することができる。
分子の安定性は、一つの分子だけでなく、幾つかの分子の組み合わさったセットとして生まれる可能性が高い。例えば、アミノ酸や核酸が重合したポリマーは私たちが考えるよりはるかに壊れやすい。溶液中のイオン構成や、あるいは他のポリマーとの相互作用により少しでも分解されにくくなれば、一つの分子単独ではなく、分子が組み合わさったセットが選択されることは十分考えられる。分子間相互作用の結果、安定性だけでなく分子の増幅ができればなお高い環境への適応性が生まれる。
実際、ランダムに形成されたポリマーの中には、新しいポリマー形成を促す触媒のような性質を持つ場合があることも知られている。例えば、塩基が10個並んだRNAポリマーがあると、さらに長い50個並んだポリマーが形成できることが示されている。他にも、アミノ酸や塩基が長いポリマーを作る段階では、できたポリマーの一部が一種の触媒の働きをして、より長いポリマーを形成するのに一役買っていることも示されている。
図2 脂肪酸膜小胞とAMP,UMPを混合して熱をかけながら、乾燥と水和を繰り返すことでRNAポリマーが形成できることを示した実験。同じような条件で、鋳型になるDNAを存在させると、10%のエラー率で複製が起こることがその後示された。
中でも興味を引くのが、脂肪酸と核酸が相互作用する状況では様々な反応の効率が上がるという実験だ。例えば、AMP,UMPのようなリン酸塩基をフォスファジル酸から作った小胞と混ぜ、熱を加えながら炭酸ガスを吹き入れ乾燥させ、その後また1mMHClを加えて溶解するというサイクルをくり返すと、脂肪膜の小胞に閉じ込められたRNAポリマーが合成できることが示された(Orig Life Evol Biosph 38:57、2008)。普通、実験室で核酸を重合させるとき、重合が自然に始まるよう核酸モノマーを活性化するのだが、この実験で使われたのはAMPとUMPで全く活性化されていない。熱と乾燥による濃縮が繰り返す条件さえ外部から与えれば、脂肪酸の助けでRNAが重合化することが示されたことは、後でのべるRNAワールドの可能性を示す重要な貢献だと考えられている。DNAの複製についてはさらに驚くべき結果が報告されている。50merの鋳型と、AMP,GMP,CMPなどをPOPA (palmitoyl-2-oleoyl-sn-glycero-3-phosphate)と混合して、熱を加えながら乾燥と水和を繰り返すと、10%程度のエラーはあるものの、鋳型に合わせたDNA複製が起こることが、2011年カリフォルニア大学サンタクルズ校のグループによって示された(Biochimei 93, 556, 2011)。以上のように、ランダムな有機分子の相互作用が脂肪膜に囲まれたRNAやDNAのポリマーの形成にまで発展できる。
このようなシステム化学の研究と、現存の生命に見られる有機高分子の機能分担を合わせて考えると、例えば複製にはDNAが適していること。一方、化学反応の触媒としてはタンパク質、脂肪が適していること。そして、RNAは両方の機能を持ち得ることや、タンパクとDNAの両方と相互作用が可能で、両者の機能を結びつけることができる分子であること。そして、必要なすべての化学エネルギーは核酸の構成要素にもなるATPを介して行われる、などの条件が整って行ったと考えられる。また、このような性質を持った有機高分子を含む分子混合物は、生命以前の地球の様々な場所に存在し、互いに反応しながら主に存在持続時間という点から選択が行われたと考えられる(図3)。
図3 原始生命誕生までの過程
しかしこれだけでは安定な個別の分子セットが選ばれるという過程の繰り返しで、発信できる情報や生物の誕生には至らない。この過程で生まれるのは脂肪酸膜で囲まれた安定で、場合により増幅可能な有機物に過ぎない。中でDNAが複製したとしても、結局は安定な分子の集合でしかない。次の段階に進むためには、有機体(生物)の語源になっている、Organize(組織化)が何らかの目的のために行われる必要がある。この目的こそが、生物が生きるという目的を示すルーツであり、また生命の基本条件になっている。この生きるという目的の一つは、生きるためのエネルギー収支を可能にする分子ネットワークで、これが何よりも最初に必要だとする考えがenergy firstモデルだ。これに対して増える・複製できることがまず必要だとするのがreplication firstモデルだ。次回から順にこの二つの考えについて見ていこう。
[ 西川 伸一 ]
熱水噴出孔と生命誕生
2015年11月16日
私たち団塊の世代が生命科学に魅せられた時必ず読めと勧められた本の1冊がシュレジンガーの「生命とは何か?」だ(図1)。

図1 シュレジンガーと著作「生命とは何か?」。版権の関係で私が読んだ岩波の「生命とは何か?」の代わりに英語版を掲載した。
生命は物理化学法則のみが支配する地球で誕生した。当然生命は物理化学法則の延長として理解すべきと考える人も多い。しかし、生命誕生によって情報も含めて地球上に新しい法則が誕生したと思っている者も少なくないと思う。私はその一人だが、そんな私にとってシュレジンガーの「生命とは何か?」は、量子力学者シュレジンガーが生命を物理学者の目で見たとき、生命固有の法則が潜んでいそうな場所を探している本に思えた。本の中で生命固有の課題として強調していたのが、生命体がエントロピーを増大させないよう、エネルギーや有機物を調達する過程と、同じ個体が増殖する過程だった。これを読んで、いずれかの問題を生物学固有の課題として考え研究を志した若者も多かったはずだ(私は途中で挫折したが)。
この分野について最近書かれた本や論文を読むと、生命誕生過程でこの二つの課題のどちらが先に起こったのかについて暑い議論が続いているようだ。もちろん独立した生物が誕生する時には両方が備わっている必要があり、両過程は卵と鶏のように互いに切り離せない。ただ素人の私からみると、生命に必須の有機物がエネルギーとともに形成されないと、最終的に核酸に頼る増殖自体も考えにくいはずで、有機物とエネルギーを合成できるシステムの誕生から考えてもいいのではないだろうか。
前回述べたが、有機物の合成の可能性については様々な可能性が議論されてきた。この中で、有機物とエネルギーを同時に持続的に生成できる場所として脚光を集めているのが、20世紀後半から相次いで存在が確認されたアルカリ性の熱水を地中深くから噴出している海底熱水噴出孔だ(図2)。この発見によって、生命誕生を有機物とエネルギーから考えるenergy firstの考え方が現在は優勢になっているように私には思える。

図2 熱水噴出孔 Wikiコモンズ
前回メタン、水素、炭酸ガス、アンモニアを熱して電気刺激を与えるとアミノ酸が生まれるユーレイとミラーの実験を紹介したが、この条件では持続的にアミノ酸が作り続けられることはない。しかも、できたアミノ酸からさらにポリペプチドを作るとなると、少なくともかなり高い濃度の有機物が持続的に供給される必要がある。反応を偶然の放電に頼るユーレイ・ミラーの条件では到底不可能だ。
ところが、熱水噴出孔には炭酸ガスだけでなく、後に述べる理由で還元力の強い水素も豊富に存在し、これらが反応してメタンやアセトンを持続的に合成することができる条件を備えている。さら地中深くから熱水が噴きあげる間に、は有機物の合成や重合化に必要な触媒となる様々な鉱物が豊富に存在すると考えられる。そして何よりも、一つの熱水噴出口は、少なくとも3万年以上との長い期間熱水を噴出し続けることがわかっており、熱水噴出孔が生命誕生に必要な有機物とエネルギーを持続的に形成できる場所であることは人の素人の私にも十分納得できる。熱水噴出孔の中には数メーターから数十メーターのチムニーという構造を形成して海水から突き出ているものが存在する。これは鍾乳石と同じで、熱水に溶けていた様々な鉱物が海水で冷やされ沈殿することで形成される。すなわちチムニーの存在は、熱水噴出が生命誕生に必要な地質学的時間維持できる可能性を示唆している。また有機物が安定に存在するのが難しい400度近い熱水もチムニーを通る間に冷却され、有機物の合成や持続の可能な温度になる。
これらの結果から、1)豊富な有機物合成の原料、2)熱や化学エネルギー、3)持続性、の点で熱水噴出孔にできたチムニーが、有機物を持続的に供給でき、生命誕生までの長期間の分子選択過程を維持できる、など熱水噴出孔が現在考えうる最適の生命誕生の現場である可能性が強く示唆される。最近、メタンを構成するアイソトープを調べた研究から、熱水噴出孔周辺に存在するメタンの少なくとも一部(1mM)は生物が関与せずに生成されたことも証明され、熱水噴出孔が有機物合成の現場である可能性は高まった。
ではどのように熱水噴出孔で有機物やエネルギーは作られるのだろう。まず炭酸ガスの還元に必要な水素だが、地殻のシリカが水と反応して蛇紋石が形成される過程で水素が発生することが知られている。水素自身はそのままだと拡散してしまうが、チムニー内に形成された鉱物の壁で隔てられた迷路のように入り組んだ小部屋は熱せられた水素を補足する。
次にこの水素により炭酸ガスが還元される過程だが、化学の苦手な私でも、水素と炭酸ガスや重炭酸塩が反応して次の化学反応が進み、
4H2+CO2→CH4+2H2O
4H2+2HCO3+H→CH3COO+2H2O
上記のように、メタンやアセトンができることは理解できる。これらの反応が起こると最終的にエネルギーは発生するのだが、反応の開始にはエネルギーを加えることが必要で、反応は自然には起こらない。従って、反応を進めるための触媒が必要になる。
このとき参考になるのが、Autotroph(化学合成独立栄養生物)と呼ばれる、無機物から自分で有機物を合成して生きている細菌類だ。メタンを合成するAutotrophは古細菌に属しており、アセトンを合成するautotrophは真性細菌のClostridiaに属していることから、有機物の原材料としてメタンを選んだか、アセトンを選んだかの偶然が、古細菌と真性細菌の誕生まで変わることなく続いたのは驚きだ。最近になってこのようなAutotrophが有機物を合成する代謝経路についての研究が進み、アセトンやメタンを合成する経路に硫化鉄を持つフェレドキシンが重要な働きを演じていることが明らかになってきた(詳細はCell, 151:1406, 2012:この原稿のほとんどはこの論文を基礎にしている)。すなわちフェロレドキシンに含まれる鉄を媒介に水素の電子を炭酸ガスに移転させる反応で、これにより炭酸ガスの還元が自発的に進行する。この結果細菌では、アセトンやメタンをATPと共に合成することができる。このフェレドキシンを媒介とする炭酸ガスの還元反応の最初に必要な自由エネルギーはアルカリになるほど低下することから、高いpHほど反応が起こりやすい。
このフェレドキシンが媒介する電子の転移をすすめる主役は鉄イオンであることから、同じ反応をチムニーに豊富に沈殿している鉄イオンで媒介できる可能性がある。アルカリ熱水噴出孔のpHは高く、水素による炭酸ガスの還元反応は起こりやすくなっているところに、フェレドキシンにも含まれる硫化鉄や硫化ニッケルが電子の転移の仲立ちをすることで、細菌で起こっている有機物合成がチムニーの中で起こるとするシナリオだ(図3)。
図3チムニー内にできた小胞内で有機物が合成される。
また細菌が炭酸ガスからフォルムアミドを合成するのに使っているモリブデンイオンも存在しており、有機物合成に必要なすべての原料が、細菌と同じようにチムニーの中で作ることができる。残念ながらこれを証明する実験はまだ行われていないが、理論的には可能だと考えられている。先に述べたように、熱水孔の周りには生物の助けなしに合成されたと考えられる1mM程度のメタンが存在していることを考えると、実験室で再現されるのも時間の問題ではないだろうか。
あとはチムニー内にできた迷路のような小胞の隔壁に存在する様々な鉱物の触媒活性を使って、アミノ酸、核酸、脂肪酸ができ始める。さらに、チムニー内に形成される熱勾配により温度拡散が誘導され、有機物が濃縮された小胞が生まれる。この中で、前回紹介したような反応が起こって有機物の重合化が起こり、鉱物膜と脂肪膜が複合した高い濃度の有機物が詰まった独立栄養系の小胞ができる。全くの素人の私にとっても、このシナリオは理解しやすく、実際に起こっているという確信を持つ。
こうして出来上がった鉱物と脂肪酸の膜で仕切られた小胞には高い濃度の様々な有機物が詰まっているが、ここまでは完全に物理化学の法則のみで進む点が重要だ。すなわち生命誕生のプロセスはここから始まる。次回はこの無機物と合体した中間段階が、チムニーからどう独立できるのか(図4)考えてみよう。
図4:チムニーないの無機物と新たに合成された有機物の合体した中間段階が、チムニーから独立した時生物が生まれる。
[ 西川 伸一 ]
分子進化をダーウィンの進化論に当てはめる
2015年12月1日
生命誕生を考えるとき、前回述べた熱水噴出孔に形成される泡のような小胞ほどよくできた偶然はないように思う。うまい具合に大きさも細胞に近く、小孔で他の小胞や外界ともつながっているし、原理的にアミノ酸、核酸、脂肪酸、ペプチド、ヌクレオチドなどを持続的に合成する条件も整っている。さらに、アセテートやメタンの合成過程でエネルギーを発生させることができ、熱勾配やpH勾配のおかげで高分子を特定のコンパートメントに濃縮することもできる。
ただ、どんなに細胞に似ていても、これは生命ではない。チムニー内の小胞では、熱力学平衡に対抗するため、不断にエネルギーを外界から導入して(ここではH2勾配、CO2の還元反応、熱などがこれに相当する)熱力学的平衡から離れた状態を維持できる開放系、すなわちイリヤ・プリゴジンが散逸構造と名付けた非平衡状態が実現できている。しかも、この状態を何万年にもわたり維持することができる。しかし、ここで起きていることの全ては生命誕生に必要な条件ではあっても、生物特有の性質とは無関係だ。非平衡の熱力学系ができただけで、生命誕生までにはまだまだ大きなギャップが存在している。一般的に創発と呼ばれてきた質的な転換が必要で、まずこのギャップで何が起こったのか大きな枠組みを考える必要がある(図1)
図1 最初の生命(LUCA:Last Universal Common Ancestor)で起こったと思われることを列挙した図。
前にも述べたが、英語では生命をOrganismと表し、生物が「組織化されている」ことを強調する。これはフランス語(Organisme)、イタリア語・ラテン語の(Organismo)も同じだ。ところが日本語やドイツ語では「生物」(Lebenswesen)と表すため、有機化学と生命誕生の間に横たわるギャップの本体が、「組織化される」過程であることを、いまひとつ実感しにくい。18世紀、デカルトの機械論では生命を理解できないと反旗を翻した自然史運動の中心ビュフォンを代表とする先人たちが目指したのが有機体論だったことを思い起こそう。その後、生物が組織化された存在だという特徴は、生物には「自然目的」が内在しているように見えるという特徴へと読み替えられ、19世紀生物学の重要なテーマとしてダーウィン進化論の誕生まで受け継がれていく。
組織化するメカニズムはそのまま訳すとオーガナイザーであり、シュペーマンが使用して以来、発生学で頻繁に用いられる用語だ。分子を生命へと組織化するという、実際には何も理解できていないメカニズムをとりあえず生命のオーガナイザーと呼んでおこう。もちろんこの本体については全く分かっていないことから、ここではオーガナイザーという言葉を、この過程を理解できていないことを表す目印として使う。と言うのも、間違うとオーガナイザーという概念は、神を始めとする超越的力による組織化と結びつけられる危険がある。事実、ダーウィン以前の生物学では、オーガナイザーとは確実に神の持つ超越的力のあらわれと考えられていたし、ダーウィン以後も「エラン・ビタール(生命力)」などの超越的力は、繰り返しオーガナイザーと結びつけられてきた。これはオーガナイザーという言葉自体に、最終目的を知り、そのために部分を組織化する主体という意味が付きまとうからだ。オーガナイザーと表現したときは、常にわかっていないことの印であることを思い起こし、超越的説明を導入しないように気をつけながら進む必要がある。
オーガナイザーを理解するため、過去の生物学の歴史にもう一度目を移してみよう。18世紀生命を組織化するオーガナイザーは、生命の「自然目的」と読み換えられていたが、この自然目的を科学的に初めて説明したのがダーウィンの進化論だった。重要なことは、進化論が超越論的説明を完全に排除してこの自然目的を説明している点だ。従って生命誕生までのオーガナイザーを考える時、ダーウィンの進化論は大いに参考になる。
ではダーウィンの進化論は生物の持つ「自然目的」をどう説明しているだろう?この説では、目的を実現すべく進化したように見える種(例えば高い木の実を食べる為に首が長くなったキリン)も、最初から小さな違いが集まった集団の中の多様性として存在し、この小さな差が他の個体と比べた特定の環境下での生殖(増殖)優位性により自然選択されただけだと説明する。すなわち、進化では目的として提示される結果が最初から存在していたという逆転の発想だ。
次回以降順に取り上げるが、ダーウィンの扱った生命誕生以後の進化と、それ以前の分子進化には様々な違いが存在する。しかし、結果が先に多様性として存在し、それが選択されるという図式は分子進化にも当てはめられるのではと考えている。すなわち、生命誕生と非平衡熱力学系の間に、無数の中間状態(分子の種類とそれらの関係性)が存在し、多様な分子の集まったカオス状態の中から、特定の中間状態が選ばれる過程が繰り返されるうちに、最後に全生物に共通の祖先(LUCA)が誕生すると考えることができる。
この分子進化のダーウィン的理解に必要なのは、1)誕生までに発生した中間段階、2)生命誕生に至るまでに必要な分子や中間段階多様性を発生させる力、そして3)分子から生命への過程に働く自然選択力になる。今回からこの3つの問題を順に考えて行く。
ダーウィンの進化論では共通祖先の概念は、出し手のいない情報を扱うための素晴らしいアイデアになっている(これについても次回以降議論する)。このおかげで、現存の生物を単一の原始細胞まで遡って構想することが可能になる。同じように分子進化でも、単純な構造(例えばディペプチドやディヌクレオチド)から複雑な構造へと分子進化が起こり、最終的にLUCAに集まったのだろうと推察はできる。もっとも単純な有機体をメタンとアセトンとすると、知る必要があるのは、このもっとも単純な有機体構造から合成されてきた、自立生命を支えるために必要なミニマムな分子の構造リストだろう。
進化論では、生物の複雑化を示す中間段階の証拠として化石の存在が指摘されている。しかし残念ながら、化石に相当する中間段階の遺物は、38億年前に起こった生命誕生までの過程を示すほど多くは残っていないだろう。再現できず、記録もない過去の分子を研究するには、現存する分子を基礎にして過去の分子を推論するための方法が必要になる。現在この目的で広く行われているのが、DNAの配列の差に基づいて系統関係を推定し、その共通祖先を推定する分子系統学だ。しかし、DNA情報が成立していない生命誕生前にはこの手法は役立たない。生命誕生までに生まれた中間状態や、分子構造の複雑化について考える分子進化は、有機分子の構造だけを指標として進化を考える必要がある。ゲノム配列を使うことに慣れた私たちには大変に思えるが、例えばダーウィンの進化論は、形態の観察だけから生まれてきたことを考えると、できない話ではない。
幸い構造解析の終わったタンパク質はすでに10万を突破している。アミノ酸配列からタンパク質の構造を自動的に予測することはまだ難しいため、タンパク質をその形態から分類しようとCATHと呼ばれる分類が1993年より行われている(図2)。
図2 Orengoらの論文で行われたタンパク構造の階層化。Cはclass, Aはarchitecture, TはTopology 。これによりタンパク質の構造の複雑化の過程を、情報ではなく、形質として研究できる。(Orengo et
al, Nucl. Acids Res. (1999) 27 (1): 275-279.より転載)
この方法では、タンパク質をClass,(類) Architecture(構築), Topology(接続形態)、そしてhomologous superfamily(相同スーパーファミリー)に階層化している。全てのタンパク質は、αヘリックス類、βシート類、そして両方が合わさったαβ類に分けられる。この3類がそれぞれ多様な変容を遂げることで異なる形態を持つ構造が生まれる。タンパク質の複雑化が進むと、同じ類に属していても、形が大きく違った構築を作ることができる。これがarchitectureだが、図1ではαβ類から派生しうる3種類のarchitectureが示されている。こうしてクラスより複雑な構造の多様なarchitectureを一定数抽出することができる。
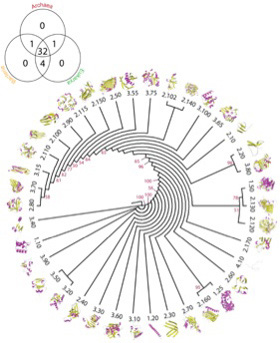
図3 Caetano-Anollesらにより提案されているarchitectureの系統図。ここでは38種類のArchitectureが区別され、構造、配列、機能を総合した推計方法で系統樹が描かれている。左端のベンダイアグラムはそれぞれのArchitectureが古細菌、原核生物、真核生物にどう分布しているかが示されている。このレベルの構造はほぼすべての生物に存在し、従ってよりLUCAの持っていたタンパク質に近い構造だと想像できる。(Bukhari and Caetano-Anolles, Plos Computational Biology,
9:e1003009, 2013より転載)
図3は、Caetano-Anollesらが、構造の決定されたタンパク質からarchitectureを抽出して系統関係を調べた図だが、重要なことはほとんどのarchitectureが古細菌、原核生物、真核生物のすべてに存在することだ。すなわち、このような構造リストは、生命誕生に必要なミニマム分子のリストに近いと考えられる。一方、様々なarchitectureが結合したtopology, homologous superfamilyなどのより高次な構造になると、存在する生物は徐々に限られていく。
このように、タンパク質やRNAの構造の階層性、細胞内での機能、そしてアミノ酸配列総合して進化過程を記述しようとするCaetano-Anollesらの努力は、生命誕生後の進化を新しい観点から見るだけにとどまらず、例えばATP合成や、アミノアシルtRNA合成酵素のような生命の基本に関わる酵素のもっとも単純な構造を推定する方法になる可能性がある。
もちろんこのもっとも単純な機能タンパクリストが完成しているわけではないが、次回からは、LUCAに集まったこの分子群のリストが完成したと仮定して、分子多様性を生み出す力、generator of diversity (GOD)について考えたいと思う。
[ 西川 伸一 ]
生命誕生までのダーウィン進化
2015年12月15日
前回、生命の共通祖先(LUCA:Last universal common ancestor)に存在すると想像される有機分子や、これら分子の祖先と考えられる様々な中間段階を、現存の生物に存在するたんぱく質やRNAの構造から推測する可能性について議論した。生命誕生までに起こった出来事の痕跡が残っていない限り、何が起こったかは現存のタンパク質から推測するしかない。もし高い確度を持つ推測方法が確立すれば、その推測に基づいて分子を再現すること自体は難しいことではない。こうして合成した有機分子の性質を調べ、最終的には生命を作り出すこともいつかは可能になるだろう。しかし、可能性としてリストされる個々の分子を実験的に検証するためには、途方もない努力と時間が必要だろう。まして、それを集めて生命を再構築することは21世紀中に出るかどうかまだわからない。だが、詳細を理解することは難しくとも、LUCAまでの道筋はぼんやりと頭に浮かぶようになってきている。
最も重要な仮定は、LUCAができる過程もダーウィン進化の法則に従うという仮定だ。他の可能性もあるかもしれないが、私にはこれ以外のアイデアはない。この過程をダーウィン進化的に述べるなら、単純な有機物から多様で複雑な中間有機物が無作為かつ連続的に生まれ(多様性の獲得)、その中から分子自体の安定性、他の分子との相互作用による安定化などによる選択が行われ(自然選択)、より安定な分子や組み合わせが選択されるといえる。この分子進化過程は、多くの偶然に左右され正確な予測はもちろんできない。しかし、基本的には熱力学的法則に従って、一種のカオスが形成される過程だと考えている。もちろん単純なカオスは生命ではない。同じ空間に共存している様々なカオス状態同士がランダムに反応し合っているうちに、次により複雑で大きなカオス状態ができる(星雲の衝突のようなイメージ参照:図1)。これが繰り返される中で、それまでにはなかった秩序が組織化されるとLUCAが生じる。

図1 双極星雲:星雲は異種のカオスだが、これが衝突して新しい一つのカオスを形成する。(写真はWikiコモンズより)
すなわち、幾つかのカオスが、力学とは別の新しいルールでOrganizeされ、これまでのカオスとは質的に異なる秩序で支配されるLUCAが生まれる。多様化と自然選択というダーウィン進化論の共通ルールは、カオスからLUCAへの過程、生命誕生後の進化過程の両方に存在しているが、両者には多くの違いがある。LUCA誕生過程を思い描くためには、この違いをしっかり理解しておくことが重要だ。
まずLUCAが誕生するまでの中間段階は、同じように自然選択されて誕生すると言っても、熱力学的法則に完全に従うカオスに過ぎない。一方、生命誕生後に生起する中間段階は、熱力学第二法則に逆らって独立した系を維持しながら増殖する生物特有のOrganizeされているという性質が最初から維持される。生命誕生後の進化ではDNAが本来持つ変異しやすいという化学的性質が、集団の多様性獲得の原動力だが、一方、生命誕生前は、多様な分子を連続的に供給する化学的仕組みが必要になる。
次に、進化の結果は生命誕生前と、誕生後では全く反対に見える。もし数少ないLUCAから生物が進化しているとすると、 LUCA誕生までの過程はLUCAへと収束する過程のように見える。一方、生命誕生後はもっぱら多様化が進んでいるように見える。図2は、生命誌研究館のシンボルとも言える生命誌絵巻だが、この扇の形は、まさにLUCA誕生後の進化過程で生物の多様化が起こっていることを表している。このことから、進化過程を洒落て、Generator of Diversity (GOD)と呼んでいる研究者もいるぐらいだ。

図2:生命誌絵巻 協力:団まりな/画:橋本律子
多様な生きものが長い時間の中で誕生した歴史と関係を表現。
詳細はこちら
実は、個体の多様化と自然選択の組み合わせでなぜ多様な種が生まれるのかについては、完全に理解できているわけではない。ただ、生命誕生後の多様性を生み出す原動力GODは、ゲノムの物質的基盤であるDNA鎖がもともと変異しやすいという物理化学的性質を持つことに起因している。おそらく祖先となる集団(共通祖先)の多様性は常に十分大きい(多様な形質をとれる)おかげで、環境により複数の形質が選択され共存するうちに種の多様化が進むのだろう(これについては生命誕生後のゲノム発生学でもう一度詳しく扱う)。生物が住める環境が通常高い許容性を持っていることが種の多様化に重要だが、寒冷化や隕石衝突のような大きな環境変化がおこると種は絶滅し多様性は減少する。
先に述べたように、一見すると生命誕生前の進化は一つあるいは少数のLUCA誕生へ向かって収束していくように見える。しかし、例えば結晶ができるように無機分子が臨界に達して一足飛びに生命が生まれるとは考えにくい。従って、生命誕生までには、比較的安定な中間段階として、多様な有機分子や分子複合体が合成される必要がある。それも、LUCAに近づくにつれ、比較的単純で多様な中間段階段階から、より複雑な中間段階が段階的に生まれていくと考えられる(図3)。こうしてできる多様で複雑化した分子集団の中から少数が選ばれ、さらに新しい秩序が生まれるよう組織化されるのがLUCAではないだろうか。従って、過程全体で見ると起こっていることは収束ではなく、進化と同じで多様化だ。ただ、強い選択圧の結果LUCAとして残るのは数少なく、収束しているように見えることになる。
図3:LUCA進化は多様化ではないように見えるが、実は分子多様化の産物だった。
最終的にLUCAが完成するにはどれほどの分子や分子ネットワークが必要か想像するしかないが、自立生命が可能になるには最低限100−300余りのタンパク質と機能的RNAが存在すればいいと想定されている。ただ、最初から無駄のない構造ができるはずはないので、おそらくこれよりはだいぶ多い分子の種類がLUCAに存在したと思える。ここでは、タンパク質とRNAを合わせて仮に500としておこう。実際には、これが100個であれ、1000個であれあまり違いはない。
ではこの500個の分子はどうすればできるのだろう?RNAについては後にRNAワールド仮説を考えるときに議論するとして、ここでは話を500種類のタンパク質を第43話で述べた熱水噴出孔で合成するという課題にしておこう。もちろん設計図などないから、ランダムな化学反応でアミノ酸が合成され、こうして合成されたアミノ酸からやはりランダムな化学反応でペプチドが合成される過程がどう進むかという課題と考えてもらっていい。
全ての有機合成が、炭酸ガスと水素から、メタンやアセトンが合成されるところから始まることを見たが、そこに窒素も加わって、様々なアミノ酸が合成できることは示されている。熱水噴出孔には触媒とエネルギーが十分存在するため、この場所ではアミノ酸から少なくとも50merのペプチドまで理論的に合成できる。この過程を図示すると、生命誌絵巻と同じで、メタンとアセトンという単純な有機物から最初始まり、時間とともに有機物の多様化が進む過程として描ける(図4)。
図4:熱水噴出孔に存在するアセトン、メタン、エネルギーを持続的に合成する力は、有機高分子の多様化のGenerator of Diversity (GOD)といえる。
このことから、LUCAへの過程も生命誕生後の進化と同じで、多様化の方向に進み、これを推進する力の基盤が、熱水噴出孔での持続的エネルギーと有機物が供給であると考えていい。最初はアセトンとメタンの2種類しかない状態が、アミノ酸になると20種類と10倍に多様化する。そしてペプチドになると、結合するアミノ酸の数に応じて多様性は指数的に増大し、もし熱水噴出孔で50merまでのペプチドが作れるなら、その多様性は20の50乗という天文学的数字になる。さらに、できた様々な長さのペプチド同士が相互作用しあって、複合体を作ることで、この多様性はさらに増大する。これは、DNAの長さが増えるごとに、それが形成できる多様性が増大するのと同じだ。このように、熱水噴出孔のような条件が整えば、GODによる有機分子の多様化を持続的に進めることができる。今後、GODが整った実験条件を工夫し、無機物から多様な有機物へと進む多様化過程を実験的に再現することは可能になるだろうと思っている。
ただ、多様化が起こったとしても、天文学的種類のペプチドが合成されるだけで何も起こらない。LUCAへと組織化されるためには、まずGODの力を何らかの形で制約することが必要だ(散逸構造の一つ翼の後ろに発生する乱気流が翼という制約により発生するのと原理的には同じだ。:図5)。

図5:散逸構造は流れのエネルギーが翼という障害物で制約されることで初めて生じる。多様化というエントロピーの流れも同じ。(写真はWikiコモンズより)
生命誕生後のダーウィン進化でも同じで、ゲノムで進むランダムな多様化を、それまでの進化の結果、課せられた生存のためのコンテクスト(それを満たせない変異が起こると個体は死ぬ)と環境への適合により2重に制約する(選択する)ことで、新しい種が生まれると説明している。
生命誕生までの過程で、熱水噴出孔に存在して、そこで発生する有機分子を多様化させる力、すなわちGODを制約する要因がOrganizerとして生命誕生を媒介したはずで、生命誕生過程を構想する鍵は、この制約要因を理解できるかどうかにかかっていると思う。もちろん理解できているというには程遠いが、熱水噴出孔にできたチムニー内に生命誕生場所を限ることができれば、実際の制約条件を特定できるのではと考えている。次回は、この制約条件を考えるときに考慮すべき幾つかの点について述べていく。
[ 西川 伸一 ]
2016年1月5日
自分で読み直しても前回の話はわかりにくかったと反省する。無生物から生物へのダーウィン進化についての思いつきを羅列しただけで、背景にある考えをよく説明できていないことが問題だった。熱水噴出孔の化学反応が原動力となって、多様な有機分子が発生する点についてはわかってもらえたと思うが、この多様化へ向かうエネルギーと物質の流れが制約されることで新しい秩序が生まれるという点については、もう少し説明が必要だろう。
有機分子やエネルギーの流れを初めから考えると複雑でわかりにくいので、ここは熱水噴出孔を一度忘れて、まず多様化と制約の一般的な関係について、もう少し簡単な系をモデルに説明してみよう。
高いところにある水は必ず低い方に流れるのと同じで、平衡に達していない状態は熱力学第二法則に従って平衡に達するまで変化を続ける。もし摩擦のない理想状態が存在すれば、この変化は川の流れと同じで、一方向に均一に流れ、新しい秩序が生まれることはない(図1上)。この流れの中央に岩のような障害物を置くと、川の流れはそれにより制約される。この結果、十分なエネルギーが供給され続ければ(川が流れ続ければ)、岩の後ろに渦という新しい構造が生ずる(図1下)。
図1 力学的エネルギーの流れが制約された時に初めて新しい秩序が生まれる。
この渦を構成する水分子を見てみると、細胞の構成成分が刻々変化するのと同じで、常に置き換わっている。しかし分子が変わっても渦の構造は、流れのエネルギーが持続的に供給される限り維持され、流れが止まると消失する。これは散逸構造の一つと呼んでいいだろう。この例では、外部の障害物がエネルギーの流れを制約しているが、制約は外部から与えられる必要はない。散逸構造として最も有名なベルナール・セル(図2)は、液体の入った容器を下から均一に熱を加えたとき、入ってきた熱エネルギーを効率よく上部へ散逸させようとして溶液中にカラム状の対流が生じて、このカラムが容器の上に美しい6角形模様として現れる現象を指す。
図2:容器の下から均一に溶液を熱すると、熱をもっとも高い効率で発散させようと、溶液分子の性質や量によって決まるサイズの対流カラムが生じる。このカラムに相当する大きさの凹みが容器上に現れる。こうしてできる構造はベルナールセルと呼ばれている(オープンアクセスの論文、Deacon and Koutroufinis, Information , 5, 404, 2014より転載)
この場合の制約は液体の粘度や比重などの性質や、溶液の量といった初期条件と言える。他にも、2種類の異なる溶液を混ぜるとき、両者が一定の法則に従って反応しあうように制約を与えると、本来なら平衡状態へと均一に混じり会おうとする2種類の液体が、周期性を持って容器内に美しい模様を描くジャポチンスキー反応がある。
図3 ジャポチンスキー反応の一例。原理は省くが、硫酸性クエン酸、臭素酸カリウム、硫酸セリウム、およびマロン酸を反応させると容器中の4価と3価のセリウム塩濃度が容器中で振動することをジャポチンスキーは発見する。溶液内の成分の反応性が制約となって、新しい秩序が生まれる好例と言える。(画像引用:https://www.flickr.com/photos/nonlin/4013035510)
図3はジャポチンスキー反応の例で、反応し合うという内的な制約だけで、これほど秩序だった模様が現れる。以上のことから、変化を続ける力学(熱力学)系に制約を加えることで新しい秩序が生まれるというイメージが理解していただけたのではないだろうか。
では熱水噴出孔での有機物生産の現場にはどのような制約が存在しているのか考えてみよう。また、この現場でダーウィン進化の選択圧となって生命に必要な分子を選択する条件も考える必要がある。ベルナール・セルやジャボチンスキー反応で生まれる構造はどんなに美しくとも、初期条件で決められた反応で、生物構造に見られる機能や目的があるわけではない。これは、自然目的が発生するために必須の、環境による選択を受けていないからだ。以上のことを念頭において、熱水噴出孔で何が起こりうるのか考えてみよう。
繰り返すが、生命が成立するためには、まず物質とエネルギーが持続的に供給される系が成立する必要がある。このような系が、水素イオンの多いアルカリ条件の熱水噴出孔には成立しているという説は説得力が高く、私もこの考えに基づいて考えている。アルカリ熱水噴出口には、炭酸ガス、水素、窒素、熱、そして反応を媒介する無機触媒が万年単位のスケールで存在し続ける。この条件がエネルギーと有機物を連続的に生成されることは化学的に不思議なことではない。この持続的Generator of Diversity (GOD)により平衡から離れた状態が維持されるが、そこで発生し散逸する物質とエネルギーの流れは、そのままだと海の中へ放出され拡散するだけだ。生命誕生の条件が整えるためには、この流れを制約する必要がある。熱水噴出孔で新しい秩序をもたらすことのできる制約の可能性をリストアップしてみよう。
熱水噴出孔から独立栄養の原始生命が生まれるまでの過程について豊富なアイデアを提案し続けているWF MartinとN.Laneの説は私のような素人にも説得力が高く、ここで展開している考えも彼らの説が核になっている。このエネルギーと物質の流れに新しい秩序を与える外的な制約として最も大事だと彼らが考えているのが、チムニー内に無数にできた、バブル状のマイクロセルだ。
図4 MartinとLaneが考えている熱水噴出孔にできたマイクロセルで進む生命誕生までの過程。この稿の多くは彼らの考えに基づいて書いている。蛇紋石化作用により水素が生成する。(Sousa et al, Phil Trans R Soc B 368: 20130088.http://dx.doi.org/10.1098/rstb.2013.0088:を改変)
これらのマイクロセルは大きさも様々で普通の細胞と同じ大きさを持つ場合も存在すると想定される。セル同士はさらに小さな孔でつながっており、生物の細胞のように独立しているわけではない。また、壁も最初は無機物からできており、その成分により有機物合成の触媒として働く。
このセルはエネルギーと分子の流れを生み出す反応容器としてまず働くが、川の流れの中の石と同じで、発生するエネギーと物質の流れの外的制約として働く。例えばフルイの役目をして、大きな分子を引き留める。これと並行して、pH、酸化還元能、そして熱の勾配もできてきた分子の制約要因として働き、熱水噴出孔のそれぞれの小部屋に存在する分子構成は、自然に不均質で多様になる。
ジャポチンスキー反応と同じで、メタン、アセトン、アンモニア、硫化水素などの成分が反応し合う法則も制約になる。加えて重要なのは、ジャポチンスキー反応と異なり、一旦合成された分子が新しくできる分子を制約する点だ。例えば現在の生物が持つアミノ酸が全てL型であるのも、炭酸ガスと水素を原料としてメタンとアセトンを合成することができるのに、メタン型とアセトン型の細菌がどちらかに収束しているも初期の制約でその後の過程が決まってしまった例だ。すなわち、小さな差でも一旦生まれると、それが増幅され維持されることで、バタフライ効果(蝶の小さな羽ばたきが最終的に大きな変化を生み出すというカオス理論)と呼ばれるものだ。
この反応の結果として生まれる制約は有機高分子になるとさらに複雑になる。例えば脂肪酸のシートができると、疎水性の面が形成される。この近傍では水が除外されるため、ペプチドが濃縮され、熱力学的に起こりにくい反応が進むようになる。また、ペプチドも水がないと分解されにくくなる。そしてすでに42話で述べたが、脂肪酸の中には核酸の重合を促進し、DNAの場合は複製酵素の役割を演じるものすら存在する。
このように、無機化学反応と比べると(例えば水素と酸素が反応して水になる)、有機化学反応は多様で、ランダムで、予想不可能だ。これは、反応自体が新しい制約要因を発生させるという、自己回帰的構造を持っているからだ。結論的に言ってしまうと、熱水噴出孔のマイクロセルでは厖大な種類の有機物の合成可能性が存在している空間ということになる。
最後に、熱水噴出孔に生まれる制約の特徴を列挙しておくと、
などをあげることができる。
最後にもう一度渦と生命誕生過程を比較して終わろう。渦の持続時間は通常短い。しかし、複雑な地形では、突然持続時間の長い渦が生まれる。もし外的な地形が変わらないとすると、渦が渦を呼び、その渦の中から安定に渦が持続する条件が生まれた結果だ。
熱水噴出孔に生まれる制約とその結果生まれる新しい秩序も安定性に大きな差が生まれる。これがダーウィン進化の自然選択に対応する。次回は生命発生に必要な状態が選択される過程について考えてみよう。
[ 西川 伸一 ]
システム化学
2016年1月15日
ここまでの話をもう一度ざっとまとめると、次のようになる。
熱水噴出孔ではエネルギーと炭酸ガス、水素、水など無機分子の持続的な流れが発生している。この流れはそのままだと、海水へと放出されるだけだが、噴出孔のマイクロチェンバーにより様々な形で外的に制約されると、有機分子の生成が始まる。最初の有機物はアセトンやメタンだが、マイクロセルの成分に含まれる様々な金属の触媒が作用すると、さらに複雑で多様な有機物が合成される。例えば以前紹介したように(第42回ゲノムの発生学II)、モンモリロン石が存在すればアミノ酸や核酸の重合が進む。脂肪酸が選択的に合成されるマイクロセルも存在するはずだ。たまたま存在した様々な外的制約により、分子やエネルギー分布の不均一化、構造化がおこる。
このようにマイクロセル内で進む有機分子の持続的な合成の流れを用いて、さらに複雑な有機高分子の合成が始まるが、ここまでのほぼ全ての過程はマイクロセルによる制約があれば進む。こうしてできた様々な分子(低分子から高分子まで)は独立したまま存在していることもあるが、高分子になるほど低いアフィニティーであっても他の分子と相互作用する確率が高まる。すなわち、合成されてきた分子自体が他の分子と反応する能力が新しい制約として働き出す。システムの中で生まれた新しい分子が、システムを新たに制約する「再帰的」制約の発生だ。
この「内的に発生する再帰的制約」について、これまでも具体的例を紹介してきた。例えば、現存するAutotroph細菌を見ると、アセトンを原料にするのは真正細菌のclostridia、メタンを原料にするのは古細菌と完全に分かれ、逆はない(第43回熱水噴出孔と生命誕生)。即ち、最初に合成された成分が制約要因としてLUCA発生から現在まで、持続的制約としてAutotrophで働き続けていることを意味する。他にも脂肪酸の中には核酸の重合や複製を触媒する能力がある分子が存在することを紹介したが(第42回ゲノムの発生学II)、これも内的に生まれた分子が、システムの新たな制約要因として働き、それ以前には存在しなかった新しい分子を作り出す分子進化の例といえるだろう。
この内的に生まれた制約による新しい状態の誕生は、制約要因が最初のシステムには全く存在しなかったという点で、反応する溶液同士を混ぜ合わせた系でおこる振動、ジャボチンスキー反応とは違う。システム内で発生した分子が制約となって、それまでとは異なる秩序が生まれるのが「内的に発生する再帰的制約」だ。
これは机上の空論だと思われるかもしれないが、応用有機化学分野のひとつ、コンビナトリアル化学では同じような課題が現実に研究されている。コンビナトリアル化学とは、例えば創薬のための候補化合物を、単純な分子から一挙に作ってしまおうという技術で、様々な化学工業で利用され始めている。従来多数の化合物を得るためには、自然に存在する生物を集め、そこから化合物ライブラリーを調整するのが普通だったが、コンビナトリアル合成のおかげで、生物に頼らず試験管の中だけで多様な化合物を合成することが可能になっている。
オランダのOttoらの総説(Li et al, J.American Chemical Society, 135:9222, 2013)は、この技術により単純な有機化合物からこれまで存在しなかった様々な新しい機能を持つ化合物が合成できることを教えてくれる(図1)。総説の内容を詳しく紹介するのは控えるが、単純な有機化合物がコンビナトリアル合成により、無限の可能性を拓くことがよくわかる。
図1 コンビナトリアルケミストリーの可能性を述べたOttoらの総説の内容を図示した。有機分子が反応を続けることで様々な複雑な高分子を合成できることを示している。
中でも私の興味を引いたのが、コンビナトリアル化学から自己複製する分子構造(複製子)が生成される可能性だ。この複製子について、この総説の著者であるオランダのSijbren Ottoのグループが昨年Nature Communicationsに発表した論文(Colomb-Delsuc M et al, 6: 7427, 2015 Creative Commons)に基づいて紹介しよう。
まずこの実験では、グリシン・ロイシン・リジン・フェニルアラニン・リジンの疎水性と親水性のアミノ酸が互い違いに繰り返すペプチドが、互いに可逆的共有結合を形成できる(SS結合)SH基を2つ持つベンゼン環に結合したオリゴペプチドが使われた(図2)
図2 Ottoらの実験では、図に示す構造を持つオリゴペプチド溶液を攪拌するだけの実験が行われた。
このオリゴペプチドはベンゼン環部分に2つあるSH基を介して他の分子と反応し、図3に示すような1merから7merの重合体を形成し、最終的な平衡状態に達すると予想される(図3)。
図3:図2に示したオリゴペプチド同士の反応から予想される分子同士の結合反応。相互に反応するベンゼン核のみを示している。
実際には7merは合成されなかったようだが、攪拌を続けると、4日目ぐらいまでに1mer,2mer,3mer,4mer,そして6merが混ざった平衡状態が生まれる(図4)。ところが、1週間を過ぎた頃から急速に他の重合体は失われ、6merが全体の90%を占めるようになる。
図4:16日間攪拌を続けた間に溶液中に存在する各重合体の比の変化。
5日目に予想された平衡状態に達するが、この平衡は1週間目に破れ、6merが急速に優位になることがわかる。
なぜ最初の平衡が破れたのかを調べると、6merの大きさの重合体だけが、今度は縦に重なって、さらに大きな重合体を形成していることが明らかになった(図5)。即ち、6merだけが縦に重なって成長する能力をたまたま持っていたため、一旦この6merが発生すると、それが核となって、6merが重合した高次構造がうまれる。この高次構造は成長に伴い、溶液中から6merは隔離するため、平衡状態が壊れる。その結果、溶液中には6mer由来の巨大分子だけが残される。
図5:6merは重なって縦に伸びる能力を持つ。この結果、溶液は6merによって占められるようになる。この重層構造は一定の大きさに達すると物理的力で壊れるが、6merの供給が続く限り、別れた部分から元の大きさの構造が形成される。すなわち、複製が起こる。
この一連の反応は、6merという構造単位の発生が新たな制約になり、これまでとは全く異なる高次構造を形成することが現実に起こりうることを示している。面白いことに、6merが縦に重なった重合体は、一定の大きさに達すると、攪拌のストレスにより、自然にちぎれる。もし6merが持続的に供給されれば、ちぎれた部分がもう一度核になって、同じ大きさを持った重合物が再形成される。この意味で、この分子構造は複製能力を持っていると言える。
このような重合による高分子形成は、ウイルス被膜や、細胞外マトリックス形成など、生命過程では普通に見られる自己組織化反応だが、全く人工的な反応として取り出してみると、生命と共通の幾つかの性質が生まれていることに気づくことができる。
これを流れの中の渦に例えると、力と水分子の定常的な供給が続く、流れの中に同じ大きさと形をした渦が増え続けるのと同じことだ。
先週Ottoたちはこの構造にグリシン・ロイシン・リジン・セリン・リジンを持つオリゴペプチドも同化できることを発見し、両方のオリゴペプチドが存在する溶液では多様な構造体が生まれることを示した(Sadownik et al, Nature Chemistryオンライン版)。この結果は、この分子構造が上の4つの性質に加えて、進化するという5番目の生命と共通の性質を持ちうることを示している。もちろん多様化だけでは進化は起こらず、特定の形質が選択されることが必要だが、このモデル系は、自然選択による進化とは何かのヒント教えてくれそうだ。
コンビナトリアル化学では最初の反応条件は人間が決める。熱水噴出孔で初期条件を決めているのは偶然だが、様々なオリゴペプチドが形成されることは既に確認されており、Ottoたちが観察したのと同じことが起こって何の不思議はない。次に、Ottoたちの研究から生命と共通するどんな性質が生まれてくるのか楽しみだ。
Otto等の研究を詳しく紹介してすこし長くなったので、今回はここまでで終わる。次回はDeaconの説を紹介しながら、生命に共通の性質の発生のためのルールについてさらに考えてみたいと思っている。
[ 西川 伸一 ]
生命誕生を考える理論的枠組み:DeaconとTeleodynamics
2016年2月1日
これまで生命誕生までに必要だと思われた過程を、なるべく具体的な例を参照しながらたどってきた。しかしここで紹介できた熱水噴出孔での化学合成過程は、生命誕生までのほんの一端に過ぎず、実際にはほぼ無数の過程がLUCA誕生に必要だったはずだ。とすると、生命に必要な様々なサブシステムが形成され、互いに連結され、LUCAの中に埋め込まれるすべての過程を洗い出すのはまず不可能だ。代わりに、この過程の背景にある一般的ルールを見つけて、より抽象的に、法則に基づいて生命誕生を理解しようとする人たちがいる。
例えばカウフマンの自己組織化理論(S.Kauffman著、米沢富美子訳「自己組織化と進化の論理—宇宙を貫く複雑系の法則」ちくま学芸文庫)、あるいはアイゲンらのハイパーサイクル理論( M.Eigen & P Schuster, The hypercycle, Die Naturwissenschaften 65:341, 1978 )もこの方向を代表する例だ。しかしこれらは生物の一部の性質を言いえているとはいえ、私たちが頭の中に描いている生物にはあまりに距離がある様に思える。
生命誕生の背景にある理論の中で、少なくとも私にとってもっとも説得力のある理論的枠組みを提供しているのがTerrence Deaconだ。今回は、彼が2006年にClayton & Davies(編)「The re-emergence of emergence:The emergentist hypothesis from science to religion」に寄稿した総説論文「Emergence:The Hole at the wheel’s hub」(http://72.52.202.216/~fenderse/Hub.htmで読むことができる)を参照しながら、生命誕生に関わる力学を見てみよう。
熱力学は、秩序のある状態と比べた時、無秩序な状態のほうがはるかに起こりやすいことを教えている。すなわち、変化のほとんどは秩序から無秩序へと進む。しかし私たちが毎日目にする様に、驚くほど多くの秩序が地球には満ち溢れている。すなわち、生命が一端誕生すると秩序は無限に増幅する。この様に生命誕生を理論的に捉えるということは、無秩序な状態(生命誕生前の)から無限に増殖を繰り返す秩序が生まれる過程に関わる理論的枠組み、この世では極めて稀にしか起こらない過程に関わる理論的枠組みを理解するということだ。秩序の増幅を目の当たりにしても、バラバラの分子からこの秩序が偶然生まれるなどと考えるのは絶対に無理だと直感的に思う。
これについてDeaconは、シュレジンガーが行ったように、熱力学法則に反する、あるいは一見法則に従わないように見える過程(一番はっきりしているのは生命)を順々に探し出し、その背景にあるルールを探している。
Deaconが徹底しているのは、「創発」、「自己組織化」、「自己触媒」など生命を表現するために使い古された言葉や概念に思考を邪魔されないよう、独自の新しい用語を積極的に導入して考えている点だ。彼の用語を用いてこれまで幾つかの例で示してきた生命誕生への中間過程を見直してみよう。
熱力学はあらゆる系に当てはまるが、生命誕生で問題になるのは化学反応が可能な溶液中の力学だと想定できる。同じ分子でも個体と液体はまったく違う物理的特性を持つ。同じ熱力学の支配下にあっても、例えば気体、液体、個体と状態が変わると、分子間の関係はより秩序の高い方向へ進む。液体ではこの分子間の関係性の反映として、例えば粘度、表面張力、熱伝導度などを考えることができる。液体中の分子の関係性は、気体への相転換が起こるまで、液体としての性質を維持し続けようとする力と考えることもできる。この液体としての分子間の関係を加味した単純な物理システムを、Deaconは熱力学と区別してhomeodynamicsと名付けている。この用語のhomeo-は、同じ関係性を維持しようとする性質がこの力学系の基本であることを表現している。この系は熱力学法則に完全に従うシステムで、エントロピーは増大し、秩序の維持に必要な制約は散逸する。
一つのhomeodynamics系は平衡へ向かって進み続ける以外何も起こらないが、この系での変化を制約すると(例えば流れの中の岩)、homeodynamicsには従わない新しい系が生まれる。これをDeaconはmorphodynamicsと呼んでいる。この用語は、ほぼイリヤ・プリゴジンの散逸構造と言い換えられ、具体的には液体が制約を受けることで発生する渦や、ベルナール・セルがこの例だ。morpho-とは形態を意味するので、形態を持たないhomeodynamicsから、渦やベルナール・セルの様な形態が発生するということをうまく表現した用語だと思う。では、homeodynamicsからmorphodynamicsが発生する背景にある共通の法則とは何か?
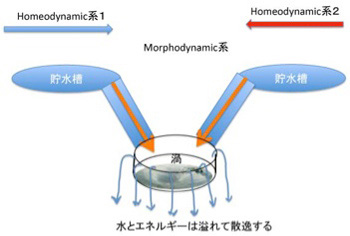
図1 morphodynamicsの発生としての渦。 2つのhomeodynamicsが衝突してカオスを形成すると、それを安定化させるために渦という形態が生じる。
もう一度渦の例でこれを考えてみよう。図1に示すように高いところにある2つの貯水槽から水を導き衝突させると、水の流れが維持される限り渦が形成される。すなわち、homeodynamicsな二つの系が逆向きに衝突して分子のhomeodynamicな動きを制約し合うとmorphodynamic 系が形成されることになる。順向きのhomeodynamic系が集まっても、エネルギーは高まるがmorphodynamic系が生まれることはない。
ではベルナール・セルはどうだろう?静置された液体の入った容器の底面を均等に熱すると、例えば固体の場合は伝導によりそのまま熱は散逸する。ところが液体の場合、表面と底面の温度差が発生すると、水分子の運動が高められ、対流が始まる。したがって、ベルナールセルを、熱の流れというhomeodynamicsと水の動きという2つのhomeodynamics系が制約し合うことで生まれるカオスを安定化させる力学として捉えることができる。すなわち、熱の流れと水分子の流れという質の異なるhomeodynamic系が制約しあっても、morphodynamics系は生まれる。
もちろんhomeodynamics系を自己組織化系と見ることもできる。重要なことは、morphodynamic系は必ず外部のhomeodynamic系が制約しあっているカオスの上に成立し、この制約が増幅することにより自己組織化が起こる点だ。
同じ様に、熱水噴出孔のマイクロセルが形成する制約による有機物生成もmorphodyamicsとして考えることができる。渦やベルナール・セルと異なり、巨視的な分子やエネルギーの流れではないが、触媒作用はエネルギーの流れを変化させる、分子の結合を促進する制約の一つだ。
ここまで見るだけでは、Deaconの考えも、生命という複雑系を、単純な力学系が相互作用してカオスが生まれるとき自己組織化が起こる系として捉える従来の考えに近い様に思える。しかし、Deaconは自己組織化やmorphodynamicsだけでは生命に必要な条件を捉えきれないと考えている。
生物はもちろん熱力学系であり、homeodynamic 系であり、自己組織化(制約化)するmorphodynamic系だが、これらをただ複雑に組み合わせても生物は発生せず、もう一段高次の力学系が必要だとDeaconは言う。この新しい力学系がteleodynamicsで、このもう一段高次の力学系を導入して生物までの道筋を整理したのがDeaconの考えの特徴と言える。
teleo-とはいうまでもなく、teleology(目的論)を意識して選んだ用語だと思う。目的、機能、複製、進化など、生物との共通性を有する増殖系として前回紹介した、Ottoの複製子を例にteleodynamicsを考えてみよう。私の理解が間違っていなければ、Ottoの複製子teleodynamics系と言っていい。
まずOttoの複製子を思い出しながら、morphodynamic系との違いを見てみよう。morphodynamic系では、分子やエネルギーの供給が続き、ミクロレベルでの分子衝突により制約が発生、拡大し、マクロレベルの秩序が発生する。しかし、分子やエネルギーの供給が止まると渦もベルナール・セルも消える。一方、アミノ酸の供給が止まり新たなOttoの複製子の合成が止まっても、複製子自体はかなりの期間存在することができ、新たなアミノ酸の供給が始まると同じ複製子の増殖を再開することができる。この差が、morphodynamicsとteleodynamicsの差と言っていい。
熱水噴出孔で実際にOttoの複製子が形成されているという証拠はないが、これが生まれるための条件を考えてみよう(図2)。すでに述べたが熱水噴出孔でエネルギーと分子の流れがぶつかり、こうして生まれるカオスが噴出孔に存在するマイクロセルの持つ様々な制約を受けてアミノ酸を含む有機分子が合成されるのはmorphodynamicsに従う過程だ。もちろん地下からの噴出が止まれば有機物は海中に拡散してmorphodynamis系は消失する。
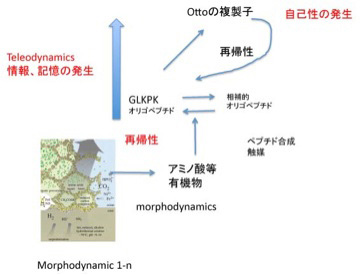
図2 Otto複製子形成過程から見たteleodynamics系の発生。説明は文中。
ここで「Otto複製子の成分GLKPKオリゴペプチドに化学的に相補的な別のペプチド(鋳型ペプチド)が存在し、GLKPKが一旦合成されると、それが鋳型になってアミノ酸重合反応を鋳型ペプチドが合成される方向へ促進する。また鋳型ペプチドは逆にOtto複製子の合成を高める」とする仮定を導入してみよう。この仮定を適用すると、GLKPKオリゴペプチドは一旦合成されると、あとは反応がこの2つの分子の合成へと制約されていく。言い換えると、有機合成が維持されるmorphodynamic系でのOttoの複製子の合成は、最初は偶然だが、一旦合成されると、同じことが安定して繰り返される秩序が生まれることになる。すなわち、GLKPKオリゴペプチドと鋳型ペプチドのどちらかが新しい制約として系内部に発生すると、GLKPKオリゴペプチドと鋳型ペプチドのセットを継続的に複製、増殖させることができる。これはmorphodynamicsでは見られなかった新しい系が生まれたことを意味している。
ここで導入した仮説はドイツの化学者Manfred Eigenが1978年の再帰的反応、ハイパーサイクルについて述べた論文(前述)で既に述べており、前回、内的に発生する再帰的制約と呼んだ系も彼のハイパーサイクルを参考にしている。
この様に、自己組織化するmorphodynamics系に、ハイパーサイクルのような再帰的関係が発生することがteleodynamicsが生まれる条件で、この関係が成立すると新しい制約を増幅させる系が現れる。もちろんハイパーサイクル自体は系として自己増殖できても化学反応にしか見えないが、前回紹介した様にGLKPKオリゴペプチドが重なり合ってOtto複製子が出来始めると、自己性を含む、生物と多くの共通点を有する力学系が発生する。Otto複製子では、複製すべき構造が環境との関わりで「情報」を発生させ、この情報に従って複製する構造が決まる。また、GLKPKオリゴペプチドはただの化学物質だが、Otto複製子の構成成分としての「機能」が生まれる。すなわちmorphodynamics系には見られない、情報、機能、自己性がteleodynamics系には存在する。このように、teleodynamics系は必ずしも生命である必要はないが、生命と共通の性質を有していることが下記らかな力学系だ。
これまで、様々な例を使ってDeaconの提唱する3つの力学系、homeodynamics, morphodynamics, teleodynamicsについて説明したが、これを図にまとめておく。Ottoの複製子を生物として見ることはできないという声が聞こえてくるが、上位の力学系が下位の力学系の単純な延長でないことは理解していただけたのではないだろうか。
Deaconの考えは、生命誕生を考えるとき、私にとってはもっとも頼りになる指針になっている。
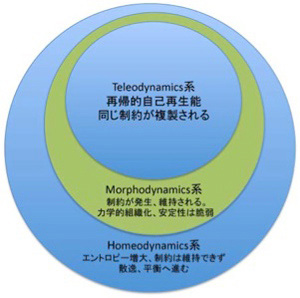
図3 Deaconの提唱する3つの力学系。それぞれの力学系は下位の系に従属する。この意味で生命も熱力学に従うが、上位の力学系は下位の力学系の単純な延長ではない。
[ 西川 伸一 ]
LUCA誕生に必要な条件
2016年2月15日
繰り返すが、Ottoの複製子(Otto Replicator: OR)は、自己性を持ち、環境との相互作用で決まる特定の構造を繰り返し複製できる点で、DeaconがTeleodynamicsと呼ぶ力学系の条件を満たしていることを述べた。しかし、だれもORを生物とは呼ばないだろう。そこで、ORと実際の生物を比較し、生命の条件を考えてみよう(図1:前回と同じ図を再掲する)。
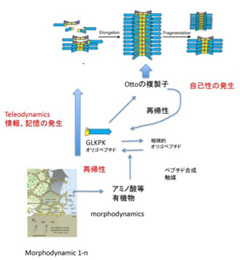
図1 前回掲載したOtto複製子についての、仮想合成経路を再掲している。これから、これと比べながら原始生命LUCAの条件を考えていく。
まずORは1種類のペプチド(図中では青と黄色の矢印)が基本単位になっているだけで、私たちが生命と呼んでいる最も単純な存在と比べても、単純すぎる。たしかに複雑だから生命と言えるわけではないが、やはり複雑性を持つことは生命の条件になるだろう。生命が複雑に見える一つの要因は、物質とエネルギーの代謝が生物には内在化しているからだ。
次にOR構造の形成過程を見てみよう。ORは熱水噴出孔に発生する(と仮定している)物質とエネルギーのmorphodynamics力学により合成されるペプチドから構成される。もちろん、OR自身も触媒や鋳型として働くことで、構築を組織化(Organize)している。しかし、生命と違って物質とエネルギーの供給は内在化されていない。一方、生物も外界からの物質に依存はしているが、エネルギーや物質代謝を生物内部で行っている。
とすると、ORは代謝を内在化させるという条件をスキップして、自己性を獲得してしまっていると言えるのかもしれない。その結果、物質とエネルギーの供給は全て外部に依存している。
このことから、少なくとも地球で見られる生命誕生の条件として、独立性・自己性が生まれるより前に、生存に必要なエネルギーや物質の代謝システムの少なくとも一部が内在化されていることを指摘できる。これがどう行われたのか想像してみよう。
前々回述べたようにMartinらはこの準備が熱水噴出孔のマイクロセル内で準備されたと考えている。マイクロセルは無機物でできており、その中での代謝システムがどれほど複雑化しても、独立することはない。その代わりに、安定した環境を長期間提供できることから、最終的に内在化することが可能な代謝システムの準備がここで起こったとする考えは説得力がある。
LUCA誕生前にエネルギーと有機物の代謝システムが生まれる過程は、現存の全ての生物に共通するシステムを参考にして考える以外にない。そして全ての生物がエネルギーと有機物合成のために共通に使っているのが、アセチルCoA、ATP、水素イオン勾配を利用したATP合成システムだ。したがって、まずこれらの分子やシステムがどう生まれたかを考える必要がある。
まずアセチルCoA,ATP分子から見ていこう。図2に示すように、アセチルCoAは、細胞内の有機合成の核として機能しており、他にも図には示していないがコレステロール合成や、ケトン体の合成など、まずほとんどの有機分子代謝経路に関わっている。おそらくわかっているものを網羅するだけで、生化学の教科書一冊分必要なはずだ(実を言うと、代謝経路はこれまで私の最も苦手な分野だ。しかし生命誕生を考えるためには必須の知識で、若い研究者はぜひ私のような苦手意識を持たないようにして欲しいと思う)。
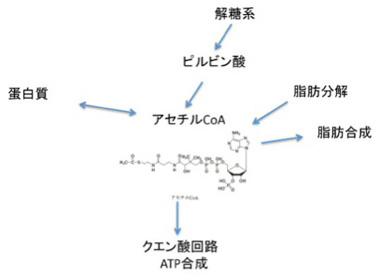
図2 アセチルCoAは有機合成の種として全生物共通に使われている。
しかし図3に示すように、熱水噴出孔では炭酸ガスが水素により還元された後、アセチルCoAを合成するWood–Ljungdahl 反応と呼ばれる合成回路が進む条件が存在したと考えられている。確かにCoAの分子構造はLUCAが生まれる条件に常に存在するにはあまりに複雑に見える。しかし、ただのメチル基がCoAの代わりに存在する極めて単純な分子でも、アセチルCoA と同じ機能を果たし得るし、またこの程度の有機分子は十分存在し得たと考えていいだろう。
図3 Wood–Ljungdahl 反応:詳細を気に止める必要はない。これまで議論した水素による炭酸ガスの還元反応からアセチルCoAの合成経路が始まることだけに注目して欲しい。(Wikiより)
アセチルCoAと同様、糖と核酸とリン酸が結合した一見複雑なATPも生物の力を借りることなく合成されることはすでに述べたので、ここではADPをリン酸化して不安定なATPに変え、それがADPに戻る時のエネルギーを化学反応に使う仕組みの発生過程について考えてみよう。
現在の生物にはATPを合成するための多様な回路が存在している。しかし、あらゆる生物に存在する仕組みは膜を隔てた水素イオンの勾配をエネルギーとしてATP合成酵素を動かしADPをリン酸化する方法だ。ただこの方法で持続的にATPを合成するためには、細胞内外の水素イオン勾配を維持する必要がある。
図4:ミトコンドリア内でのATP 合成のメカニズム。説明は本文参照(Wikiより)
私たちの細胞でATP合成を行なっているミトコンドリアでの仕組みを見てみよう(図Wikipediaより)。電子を移送するトランスポーターが膜上に並んでおり(図にはI,II,IIIとナンバーを付けて示している)、電子が伝達システムの分子間を移動するたびに水素イオンがミトコンドリア外に放出される様にできており、これによりミトコンドリア内に流入した水素イオンは細胞質へと汲み出される。こうして維持される水素イオンの勾配を使って水素イオンはミトコンドリア膜上にあるATP合成構成酵素を通って流入する。このエネルギーを用いてこの酵素はATP合成を行う。(より詳しく知りたい方は、季刊生命誌43号の図もご参考ください)
ATPを作るために水素イオン勾配を使うシステムはあらゆる生物に見られるが、エネルギーのために常に流入する水素イオンをくみ出して勾配を維持するためには、このように高度に組織された複雑な構造を持つ分子セットが必要になる。もちろん、LUCAが誕生する最初からこのような分子が存在するはずはない。しかし、水素イオンの勾配を維持する仕組みがないとエネルギーを自分で作れないというジレンマに陥る。実際、薬剤でこの電子伝達系を止めると、ATPの作れなくなった細胞は即座に死滅する。
どうすれば複雑な電子伝達系なしに水素イオンの勾配を維持すれば良いのか?この問題を解決する一つの素晴らしいアイデアが、Nick Laneらにより示されている(Sojo et al, PlosBiology 12: e1001926, 2014:図4)ので紹介しよう。このアイデアは、特別なメカニズムなしに水素イオン勾配が利用できる方法を示すだけでなく、生物の自己化、独立化の過程も説明できる可能性を秘めたアイデアだ。
図5 Laneはアルカリ熱水が地下から噴出するベントでは、水素イオン勾配が維持できる条件があると提唱している。
LaneはMartinとともに、熱水噴出孔でLUCA誕生の準備が進んだと考えている一人だ。マイクロセルは小さな穴が空いているが、分子透過性の選択性はない。図では、この穴に水素イオンが透過する脂肪膜を持った小胞が詰まるという状況を設定している。熱水噴出孔では、地底から湧き出すアルカリ性の液体と、弱酸性の海水が存在するが、それぞれの流れが脂肪胞とマイクロセルを形成する無機物で隔てられると、持続する水素イオンの勾配が図の上下で生まれる。地底からの液体と、海水は常に流れているので上下のpHの差は維持され、水素イオンの勾配も維持される。
ここに図5・緑色で示した水素イオンの流れをATP合成に使える原始的タンパク質が誕生すれば、水素イオンを脂肪胞外へ汲み出す複雑なメカニズムが存在しなくとも、ATP合成システムを持つ脂肪小胞が生まれる。その後、同じ脂肪胞内で水素イオンと他のイオンを交換的に移行させるトランスポーター分子がたまたま合成されると、水素イオンの汲み出し機構とATP合成システムの両方を備えた脂肪胞が完成する。
このシナリオが実際に起こったかどうかはわからない。しかしこの考えは、LUCAに必須のATP合成システムとアセチルCoAを併せ持った脂肪胞の形成過程を、大きな飛躍をせずに考えられることを示唆している。少なくとも私は、エネルギーと有機物質合成系を備えた脂肪胞が熱水噴出孔で発生することがそれほど困難でないと確信できる。また、このような細胞様システムを人工的に合成して研究が行われることも遠くない話だと思っている。
とはいえ、エネルギーと有機物の合成系を内在する小胞を先行させて考えるとき、次にもっとも難しい問題として登場するのが複製だ。ORのような単純な構造の複製は容易だが、系が複雑になればなるほど、その複製は困難になる。
有機物とエネルギー合成システムを備えた細胞様小胞の実現性を確認した後で、十分複雑化したシステムにどうすれば複製能力を付与できるかを考えていこう
[ 西川 伸一 ]
ダーウィン進化自体が情報を発生させる
2016年3月1日
前回までのLUCAへの道(個人的に可能性が高いと考えている道にすぎないことを断っておく)を手短にまとめると、次のようになるだろう。おそらく水素と炭酸ガスから、アセチルーCoAあるいは、より単純な代換え化合物を合成するためのWood-Ljungdahl経路が完成する(49話図3参照)。これが脂肪膜内で起こるのか、原始のスープの中で起こるのかはわからない。しかしこれが完成すると、様々な有機物を合成できるようになる。もちろん脂肪膜も形成される。こうしてできた脂肪膜は小胞を形成するだけでなく、強力な触媒としても作用することはすでに述べた。アセチルCoAと一定の触媒機能を備えた脂肪膜小胞が比較的高い確率で持続的に形成され、たまたま自然に形成された水素イオン勾配の存在するマイクロセルに引っかかっているうち、水素イオン勾配をATP合成に使える小胞が生まれてくる(ATP合成酵素の誕生)。ここまでが前回述べたシナリオだ。
しかしこれだけでは自然の水素勾配が存在する場所から離れることはできない。独立したATP合成系を持つためには、細胞膜内外のプロトン勾配を維持するメカニズムが必要になる。もちろんミトコンドリアにあるような電子伝達系は複雑すぎて、それが備わるのはずっと後の話だ。この最も単純なプロトン勾配維持システムについてもNick Laneの仮説は十分説得力があるので紹介しておこう( Sojo et al, PlosBiology 12: e1001926, 2014)
図1 独立したATP合成系を備えた脂肪小胞の形成過程(Sojo
et al, PlosBiology 12: e1001926, 2014より)。説明は本文参照。
自然のプロトン勾配を利用するATP合成酵素ができるところまで(図1Aまで)は前回紹介した。この仮説では、次の過程をプロトンとナトリウムイオンを反対の方向に移動させるアンチポートの誕生としている(図1B)。膜の透過性がザルのように高いとき、このアンチポートの機能的意味はないが、膜がナトリウムを通さなくなると、プロトン流入の度にナトリウムイオンを汲み出すメカニズムとして働くことができる。この分子に加えて、さらに小胞中からナトリウムの汲み出しを高めるために、プロトンとナトリウムのどちらでも汲み出せるエネルギー依存的ポンプができると(図1C)、プロトン勾配を維持するための準備が整う。プロトン通過性の膜を持つ小胞の場合、この分子はもっぱらナトリウム汲み出しに使われるが(図1D)、プロトンを通さない膜に変化するとプロトンを汲み出してプロトン勾配を維持するメカニズムとして働くことができる(図1E)。このようにして独立したプロトン勾配維持システムが出来上がると、外界の条件にとらわれずATP合成に利用する小胞が完成する(図1)。
さて、ここまで順を追ってNick LaneやWilliam Martinの考えを中心に、ありそうなシナリオをつないでみると、読者の皆さんも独立した代謝システムを持つ脂肪胞なら十分合成できそうだと納得されるのではないだろうか。しかし本当は絵に描いたようなシナリオに簡単に納得しないでほしいと思っている。実際に重要なのは可能性のあるシナリオではなく、どのシナリオであっても、その過程が繰り返し起こる条件を明示できるかどうかだ。
ダーウィン進化では、特定の形質に情報、すなわちゲノムが対応し、環境にフィットした性質を生殖を通して(すなわちゲノムの複製により)増幅することができる。しかし、ゲノムが発生する前にはこの手は使えない。
ゲノムが存在しない時のダーウィン型進化とは何かをまず考えてみよう。生物が介在しない有機合成系では、炭酸ガスや水素といった単純な無機物から、複雑で多様な有機物が持続的に発生している。ただこの合成はランダムで、基本的には再帰性はない。従って多様性がますます増大する、すなわちエントロピーが増大するhomeodynamic過程と考えられる。この中から、例えばATP合成系を持った脂肪小胞が生まれるということは、エントロピーが増大するよう一方向に進む反応系を制約して、特定の秩序が自然選択される条件が存在することになる(図2)。
では何が選択されるか。もっとも理解しやすいのは、分子の安定性を指標とする選択だろう。普通有機物の寿命は短い。しかし、環境条件にフィットした分子は長い時間存在することができる。あるいは温度勾配を利用したふるいにかかり、特定の大きさの分子が特定のマイクロセルに濃縮され選択されることでも多様性を減じることができる。さらにはマイクロセルを構成する無機質の持つ触媒効果も当然多様性の制約に働く。これらが合わさって特定の構造が安定に存在できると、合成自体はランダムであっても、自然選択により系内の多様性を減少させることがある程度可能になる。
図2 分子進化における外界による自然選択:多様な有機分子を様々な外的要因で選択して、特定の構造の合成を繰り返して起こすことは可能か?
注意して欲しいのは、環境要因が多様な分子集合の多様性を減らす制約として働くとき、巧まずして一種の情報が発生していることだ。すなわち、こうあるべきという方向性を環境が与えることで、存在可能な分子の種類を制約している。ただ、できてきた分子を環境によって選択するだけの情報では、制約の程度は知れており、似た種類の分子の濃度は上昇するかもしれないが、同じものが繰り返し合成されるほどの情報にはならず、これだけでは情報は萌芽段階にとどまる。
では次に持続的な情報が発生するには何が必要か?
ここでもう一度コンビナトリアル化学で紹介したOttoの複製子が生まれる過程を思い起こしてほしい。
図3 Otto複製子の発生過程 説明は本文。
重要な実験なので図3に再掲するが、Ottoの複製子はGLKFKと5種類のアミノ酸がSH基を2個もつベンゼン環に連なっている分子を試験管内で反応させているうち、突然現れる構造だ。最初はSH基同士が反応重合して多様な化合物を作るが(図3b及びd)、ある時突然多様性が失われ、基本構造が6個環状に結合した構造(6mer)だけに収束する(図3d)。これは6merが発生し、6mer同士が今度は縦に重合し始めると、それ自身が反応を6merへ収束させる能力を持つからだ。すなわち、合成物自体が制約として、合成反応自体を制御するようになると、ランダムな反応が特定の構造に収束することがあり得ることを示している。
この制約は物理学の法則とは全く異なる。すなわち、反応は最初に生まれた再帰的制約能力を持つ分子の性質によって決まり、もう一度最初から反応を繰り返した時、同じ制約が繰り返して現れる保証はない。特に最終産物が複雑であればあるほどその確率は低い。物理法則とは違い、偶然に支配されているが、一旦発生すると反応が向かうべき方向性を情報として指示するようになる。
図4 発生した合成物が反応自体を制約することができると、反応の方向性を決める情報が発生し、制約された分子集団が安定して合成されるようになる。説明文中。
原理的には、ここで説明したのと同じような情報の発生と、それに制約された過程が、LUCA誕生過程でも複雑な形で起こったのだろう。これまで見てきたように脂肪膜や、アセチルCoAなどは再帰的制約を形成する能力が高い分子だ。実際には何段階にもわたって制約=情報が発生し、より生命に近いteleodynamics系が発生したと考えられる。
しかしこうして比較的複雑な細胞様構造が発生し、この構造が、合成の方向性を決める一種の情報による制約により繰り返し合成できるとしても、合成物自体と環境がリンクして自然発生した情報は、情報としては生命誕生には十分でなかったようだ。すなわち、現在の生物には独立し系全体の複製をリードできる情報が存在している。この情報の独立が次のステップだ。このステップにおそらくRNAワールドと呼ばれている系が必要になる。次回はこのRNAワールドについてみることにする。
[ 西川 伸一 ]
情報の独立
2016年3月15日
Ottoの複製子をモデルに、ランダムな化学合成系の過程から、この反応を制約する分子が誕生すると、それが情報と同じ働きをして、反応を一定の方向に導き、一定の構造を複製し続けることを前回示した。これほど強い制約でなくとも、多様化する有機分子が環境条件によりダーウィン的選択をうけ、特定の構造が濃縮されるよう制約されること自体も、考えようによっては、情報が発生していると見ることができる。繰り返すがここでいう制約は物理法則とは違って偶然発生したもので、この制約に基づく情報は独立して存在しているわけではない。すなわち少しでも環境が変化すれば、その情報は消失する。従って、ここではこの様な情報を偽情報と呼んでおく。
偽情報だけでどこまで複雑なシステムが形成可能か?計算機によるシミュレーション実験が必要だろうが、偽情報だけで達成できる複雑性には限界があると思う。これが生命誕生以前の有機合成システムがもつ物理的限界になる。
例えば前回述べたATPエネルギー合成系と、有機分子合成系を両方備えた程度の複雑性をもつ脂肪胞の複製でも、偽情報だけでは安定に複製することは到底不可能ではないかと考えている。では次の段階として何が必要か?
この答えを地球上に存在する生物に求めるとすると、「情報がゲノムとして独立すること」と言っていいだろう。現存のあらゆる生物はセントラルドグマに従い、DNAを媒体としてゲノム情報を独立させ、この情報を複製により増殖するとともに、転写により細胞の維持に使っている。そして、生物が介在しない有機合成系では到底到達できない複雑性を達成している。
ただ、自律的に有機物とエネルギーを形成する代謝システムが、一足飛びに現存の生物が持つ、独立した情報を中心にしたシステムへと転換することは至難の業で、幾つかの中間段階を介在させて考える必要がある。この中間段階として提案されているのが、いわゆるRNAワールド仮説だ。この仮説には様々なバージョンがあり、エネルギーや物質代謝システムに先行してRNAワールドが存在したと考えるreplication first仮説もあるが、ここでは最低限の独立代謝システムと並行して、あるいはその成立後に加わる系として考えていく。
もちろん、どちらが実際に起こったのか、あるいは本当にRNAワールドが存在したのかに答えることは現段階では難しい。ただRNAはそれ自身で明確な情報と機能が共存する有機システムを形成する能力を有し、LUCAへの中間段階として働く可能性がある。まず、その点について見てみよう。
RNAワールド仮説は、タンパク質と同じようにRNAがリボザイムと呼べる様々な酵素活性を持ちうるという発見が基礎となっている。もともとRNAはヘアピンループ構造を基盤とする複雑な立体構造を取り得ることが知られており、タンパク質と同じような酵素活性を持てるのではと期待されていた。1980年に入ってすぐ、後にノーベル賞を受賞するThomas CechとSidney Altmanが、スプライシングや前駆体tRNAが機能をもつ成熟tRNAになる際のプロセッシングにRNAが酵素として働いていることを発見する(図1)。
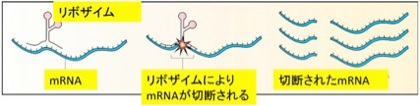
図1 RNAからできたリボザイムによるmRNA切断を説明したウィキメディアの図(https://commons.wikimedia.org/wiki/File:Ribozyme.jpg)。Thomas Cechはリボザイムを含む一本のmRNAが、タンパク質の助けを借りることなく自分自身を切断融合する(スプライス)ことを示してノーベル賞を受賞した。
それ以後研究は急速に進み、現在リボザイムは様々な生体機能を調節するために有望な分子として創薬などに利用され始めている。例えば50merほどのRNAをランダムに合成して、その中から特定の分子と結合するRNAを生成して、阻害剤として使うというような使い方が行われている。なぜ期待が大きいかというと、活性のあるRNAを容易に合成できること、それ自身が塩基配列情報を持っているため、同じ分子を複製することが容易であるためだ。
このようにCechとAltmanの発見により、それ自身についての明確な情報を担ったRNAが同時に、酵素活性という機能を持ち得ることが明らかになった。すなわち、偽情報と比べた時、RNAワールドでは、情報が独立性を持っている。以前述べたが、情報は物質ではない。リボザイムではRNAという媒体で表現されていても、次の段階でDNAということなる媒体で同じ内容を表現することができる。まさに、DNAを媒体として独立した情報を持つLUCAの中間段階にぴったりの存在であることがわかる。
ただいくらRNAが情報としての側面と、機能分子としての側面を持つと言っても、最初はRNAが情報に基づいて合成されることはない。少しわかりにくいが、この段階ではランダムにRNAが合成される。すなわち外で降る雨と同じで、自然現象と言える。ただ、できた情報を何らかの方法で解釈することができるようになると、ここに情報が誕生する。ランダムに自然現象として生まれたRNAの中に、このRNAが持つ情報を解釈するRNAが誕生したと考えるのがRNAワールド仮説だ。
これまで説明してきたように、条件が整えば自然現象としてヌクレオチドを合成することができる。またフォスファチジルコリンを主成分とする脂肪小胞はRNAを重合させる活性が高く、50merまでのランダムなポリヌクレオチドを合成するための触媒活性を持つ(Orig Life Evol Biosph 38:57、2008)。従って、現在研究者がリボザイム合成として行っていることは、自然現象として起こる。研究者は、自分の目的に合わせてこの中から特定のRNAを選択し、そこにある情報を使うが、RNAワールド誕生では、まず誕生したRNAの中に、RNAの塩基配列を解釈できる機能が生まれることが必要になる。従って、一般的にRNAワールド仮説ではRNA配列を解釈して複製するリボザイムの誕生を最初のゴールと設定している。
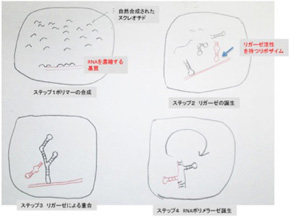
図2 RNAワールド誕生までの過程を描いたJoyceらの仮説。
(Cold Spring Harb Perspect Biol 2012;4: a003608より改変)
Joyceらは先に述べたRNAの重合を触媒する脂肪膜と、RNAが結合して濃縮される基質を備えた小胞の中でRNAの分子進化が進み、RNAポリメラーゼが誕生するまでの過程を図2で示すように描いている。まず、ランダムにポリマー化したRNAはヘアピンループ形成を介して、多様な構造を形成する。ただ、ランダムに合成されるRNAの長さには限界があるので、さらに複雑なRNAを合成するためにはまずRNA鎖同士を結合するリガーゼ活性を持つリボザイムが発生する必要がある。この結果ヘアピン同士が重合を始め、より複雑な構造の大きなRNA分子が形成される。こうして生まれる分子構造の中にRNAポリメラーゼ活性を持つ分子が誕生することでRNAワールドが完成することになる。一旦RNAポリメラーゼが誕生すると、小胞の中に存在するRNAを鋳型として複製が行われる。もちろんポリメラーゼ自身の複製もおこる。
このシナリオを実験的に証明することは難しい。すなわち最初はヌクレオチドしか存在しない状態からスタートして、RNAの分子進化を介してRNAポリメラーゼを誕生させ、自己複製が可能なRNAワールドを形成する実験は行われていない。しかし自己複製可能なRNAシステムが可能なことは理論的にも、実験的にも示されている。
繰り返すがRNAワールドの誕生は、自然に合成されているときは情報でも何でもないRNAの塩基配列の中に、塩基配列を解釈できるリボザイムが誕生した時点で始まる。この時、それまで情報でなかったRNAの塩基配列が、情報に変換される。そしてこうしてできた情報を増殖させられる系は、増殖できず一回で消滅する情報と比べると、当然自然選択の対象になる。また、情報のエラー発生を通して、情報は多様化する。解釈するリボザイムが存在する限り、エラーが起こった情報も、情報として処理され、自然選択を通してより環境にフィットした系が進化することになる。ただ、このRNAワールドは、RNAだけで完結している。従って次のステップは、多様化と自然選択が可能なRNAワールドが、例えば独立代謝システムと相互作用し、両者が組織化された系の誕生だ。次回は、この過程について考えてみる。
[ 西川 伸一 ]
参照情報と記号情報の誕生
2016年4月1日
前回、RNAワールドでは、自分を鋳型として新しい分子を複製できるシステムが可能になっていることを説明した。同じような現象は、プリオンなどでも見られるため、RNAワールド以外でこれが起こらないことを証明することは難しい。もちろん私はこの分野の素人だが、それでもRNAがランダムに合成されているうちに、リガーゼやリプリカーゼ活性のあるリボザイムが生まれ、それによりRNA自体が情報として独立するとする考えは理解しやすい。また、ここで議論しているRNAの特徴を利用して、アプタマーなどRNAテクノロジーが進展しているのをみると、特定の機能が塩基の配列として解釈され、塩基配列から生まれる機能分子を複製することが可能なRNAワールド仮説は極めて魅力的だ。
また分子の機能を塩基の配列として解釈し複製できることは、RNAワールドでは情報が環境から独立したことを示す。例えばOttoの複製子も同じ分子を複製するが、この時の情報は前にも述べた偽情報で、複製子自体の物理化学的性質と環境条件によって形成されており、環境が変わると、情報の内容も変化する。一方RNAワールドでの情報は、もちろん大きな意味で物理化学法則(RNAという媒体を使う点)に従い、条件によっては情報として読み出せないこともあるかもしれないが、原則として塩基の並びとして表現されており、周りの条件からは独立できている。
このようにRNAワールド仮説は、機能を周りの条件から独立した情報として表現できることを示してくれたが、この情報はRNAワールド内だけでしか働かない。現在のような、アミノ酸、タンパク質、リボゾームRNA、mRNAなどが入り混じったセントラルドグマが成立するためには、まず特定のRNA分子がアミノ酸と対応する参照関係やコドンとアミノ酸の対応のような記号化が必要になる。現存の生物を眺める時、アミノ酸との参照関係と記号化(コドン化)が最もはっきりと見られるのがトランスファーRNA(tRNA)だ。次に、tRNAとアミノ酸との参照関係がどのように生まれたのか、またtRNAが塩基配列を記号として認識できるようになった過程を見てみよう。
まず大学生時代に戻って、トランスファーRNAとはどんな分子だったか思い出してみよう。
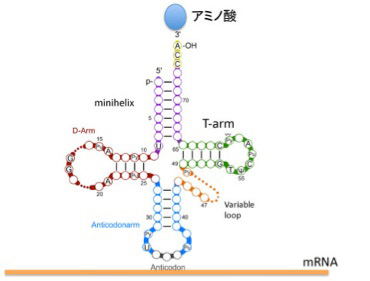
図1 tRNAの構造。ミニヘリックスとT-ArmはアミノアシルtRNA合成酵素が認識する。一方、D-ArmとAnticodonarmはリボゾームと相互作用する。(Wikicommonsを改変)
図1に tRNA共通の構造を示すが、平均76塩基からなるRNAで、ミニヘリックスを形成する3’末端に特定のアミノ酸が結合し、アンチコドンループでmRNAと結合する。この構造から、tRNAがアミノ酸と核酸の参照関係を成立させている分子であることがよくわかる。すなわち、アミノ酸とtRNA自身の対応関係を、mRNA、そして最後にはDNA上の塩基配列へと拡大する役割を担っている(後述のように実際には簡単ではない)。
このtRNA自身と、アミノ酸との参照関係の成立には、各アミノ酸に対応して存在するtRNAを認識して、それに対応したアミノ酸を結合させるアミノアシルtRNA合成酵素が関わっている。
図2に示すように、アミノアシルtRNA合成酵素(aatRNA合成酵素)は、1)ATPによりアミノ酸を活性型に変え、2)そのアミノ酸に対応したtRNAと結合して、3)アミノ酸を3’末端に共有結合させることで、tRNAとアミノ酸の参照関係を成立させる。すなわち、各tRNAとアミノ酸の特異的な参照関係を決めているのは、まさにこのaatRNA合成酵素ということになる。しかし、こんな都合のいい酵素は最初から存在するはずもなく、アミノ酸と特定のRNAの対応関係を成立させるための単純で自律的なシステムを構想しなければならない。1982年これを可能にするシステムについて清水幹夫はC4N仮説を提唱した。
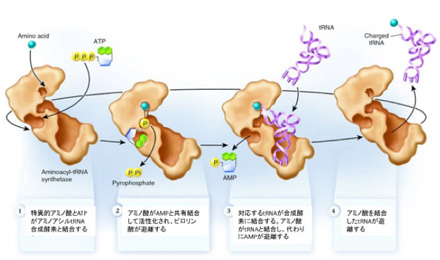
図2: アミノアシルtRNA合成酵素によるアミノ酸とtRNAの結合プロセス(Wikipediaを改変)
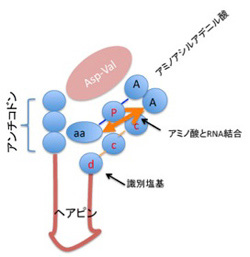
図3 清水幹夫の論文(J.Biochem. 117, 23-26, 1995)に書かれていたC4N仮説が想定している、原始型tRNA.
図3は清水の1995年の論文(J.Biochem. 117, 23-26, 1995)を改変したものだが、tRNAのアンチコドン部を5’末端に持ち、現存のtRNAのミニヘリックス部分に似たヘアピン、識別塩基、CCA3’末端と続く短いRNA分子(例としてグリシンに対してはGCCGGCGAUGUUCGCAUCGCCUCCA)が、アスパラギンとバリンからできたペプチドと結合すると、特定のアミノ酸(この場合グリシン)に対応した特異的ポケットを作り、Asp-Valの触媒作用でアミノアシル化されたRNAに変換できるという仮説だ。1995年の論文では、合成したRNA、アミノアシル化グリシンアデニル酸、そしてAsp-Valを反応させると、アミノ酸がRNAに転移できることを示している。さらに、アンチコドン部分をアラニン(AGC),バリン(GAC)に変更すると、今度はそれぞれのアミノ酸が特異的にRNAと結合することから、アンチコドンが3’末端の識別塩基と協調することで特異性を発揮できることも示している。すなわち、アミノ酸ダイマーが共存しておれば、RNAが自律的にアミノ酸との参照関係を成立できる可能性を明らかにした画期的な論文といえる。
もともと、Gly-Glyのようなアミノ酸のダイマーは、RNAの分解を阻害する能力があることが知られていた。したがって、アミノ酸ダイマーと特定のRNAが安定的な組み合わせとして選択されていた可能性は十分考えられる。ただ残念なのは、2013年の東京理科大学の田村さんの総説を読むと(Viva Origino 41, 29-33, 2013)、「C4Nは非常に画期的なアイデアであるが、今後、この仮説を実験的に検証していく必要がある」と書かれており、1995年の実験がまだ再現されていないことをうかがわせる。とはいえ、C4N仮説は一石二鳥のアイデアで、RNAワールドに、アミノ酸への参照関係性を成立させ、この参照関係をコドンとして記号化することまで視野に入れた独創的な仮説だと思う。
ただ、ここまでうまく話が進まなくとも、NCCA配列を3’末端に持つRNAが、それ自身で、あるいは他のリボザイムの助けを借りてアミノアシルRNAを合成することは十分可能だろう。一方アミノ酸との参照関係を記号化するコドンの誕生については、アミノ酸自体の化学的性質と、塩基配列で表現されるコドンの間に物理化学的・自律的な対応関係が存在しており、少なくとも一部のアミノ酸に対するコドンルールの誕生は化学的に説明できると考える人は多い。
ただ、tRNAに存在するアンチコドンは、アミノ酸の並びを指示するmRNAなどの指令が存在しなければ意味を持たない。すなわち、アミノ酸とRNAワールドをつなぐ媒体はできたとしても、この媒体を使って指示を行う情報はまだ全く存在していない。では、原始tRNAは何をしていたのか?
tRNAの原始型はアミノ酸を濃縮するために誕生してきたのかもしれない。生命誕生前でも、アミノ酸が生成し、その重合が起こりうることは示されているが、重合するにはアミノ酸の濃度が一定以上である必要がある。しかし、実際にはランダムに作られるアミノ酸濃度を高く保つのは難しかっただろう。したがって、アミノ酸を一定の場所に拾い集めることが重合の条件になり、この役割を原始tRNAは担っていたのかもしれない。。言い換えると、原始型tRNAの持つアンチコドンはあくまでも特異的アミノ酸を拾うための仕組みで、アミノ酸をくわえたRNAがヘアピン構造を介して特定のマトリックスに結合し、ランダムではあってもアミノ酸を特定の場所に濃縮することで、ペプチド合成の可能性を高めていたと考えても不思議はない。すでに述べたが、アミノ酸ダイマーの一部はRNAを分解から保護する作用があり、その合成を促進する系は進化的優位性がある。
こう考えると、図3のC4N仮説でヘアピン構造として示されているアームは、ひょっとしたら、マトリックスに結合するためのアームとして使われたのかもしれない。そしてこのマトリックスがDNAであった可能性も十分ある。おそらく生命誕生前に最も多く存在した高分子の一つがATPだろうが、それが重合すれば安定度抜群のマトリックスDNA鎖ができる。また、DNAはRNAともタンパク質とも相互作用可能な高分子だ。アミノアシル化されたRNAを濃縮して、ペプチドを合成するには最適の高分子と言っていいだろう。今述べたのはただの妄想だが、DNAとRNAの関係はこんなところから始まったのかもしれない(図4)。
図4:DNAをマトリックスとして、原始型tRNAが集合してペプチドの合成が促進する可能性を描いた図。
この段階では(図4)、アミノ酸と特定のRNA分子が参照関係を形成し、(C4N仮説が正しいなら)、さらにアミノ酸とコドンとの関係まで成立していたためかもしれないが、まだtRNA上のアンチコドンと対応させる情報はどこにも存在しない。この場合、ペプチドはあくまでもランダムに作られる。こうしてランダムに合成されたペプチドを環境との適応性を指標に選択したとしても、その選択結果を記録する逆セントラルドグマが可能であることを示す証拠は現在のところ見当たらない。
いよいよ問題は核心に入ってきたが、最後に残る課題はこの段階から、DNAへの情報の移転がどう起こったのかを説明することになる。
この過程に関してはおそらく2つの説明が可能だろう。一つは、選択されたペプチドのアミノ酸配列を記号として解釈する方法が存在するとする考え方と、アミノ酸を濃縮するためのマトリックスとして使われてきたDNAが非特異的マトリックスから、コドンでアミノ酸と対応する媒体へと変換をはたし、このランダムな配列から合成されたペプチドの選択を通して、DNA配列が選択され、遺伝子が生まれるとする考えだ。
この問題をさらに掘り下げるため、次回はアミノ酸の重合を支えるリボゾームRNAについて見て行こう。
[ 西川 伸一 ]
独立した遺伝情報誕生
2016年4月15日
生命誕生以前、コドンやアンチコドンに対応する塩基配列は、コードとしてではなく、その物理化学的性質に基づいてアミノ酸と化学的に結合していた可能性を前回述べた。さらにtRNAの中には、アミノ酸が結合したアミノアシル化tRNAを合成するリボザイム活性を持つものが存在することも示されている。従って、原始tRNAは、酵素の力を借りることなく特異的アミノ酸をキャッチし、そのアミノ酸を3’末端に結合させたアミノアシルtRNA(aatRNA)を合成できる。
例えば前回述べた清水のC4N仮説によると、この時現在のアンチコドン塩基配列がアミノ酸をキャッチするのに使われるが、この使い方は、現存のtRNAのように、コドン/アンチコドンのようなコードとしてではない。くどくなるのを恐れず、繰り返すと、原始tRNAにたまたま現在のアンチコドンと一致する塩基の配列が存在していても、アミノ酸との対応が記号化した情報として表現されているわけではない。アンチコドンとアミノ酸が確実に対応づけられていても、これはあくまでもアンチコドンの持つ物理化学的性質の反映だ。
やはり前回、コドンを持つmRNAが存在しなくても、アミノアシル化したtRNAが何かのマトリックスを利用して集まることができること、また濃度の低いアミノ酸をキャッチして一定の場所に濃縮するためにtRNAが誕生したのではないかと述べた。こうしてアミノ酸を濃縮する事でポリペプチドへと重合させる可能性が高まる(52話の図4を再掲)。即ち、RNAとペプチドの基本関係(物理的)がこうしてできあがる。
図1:52話の図4を再掲している。今回の図では、ペプチド形成は、原始リボゾームのペプチド転移活性によることを明確に記載している。
この時、tRNAに結合したアミノ酸をポリペプチドへ重合するペプチド転移酵素が必要だが、RNAワールドではもちろんこれを行っているのはリボザイムだ。現存のリボゾームはRNAと約50種類のタンパク質が結合してできているが、ペプチドを結合する活性を持つのは純粋にRNAから出来たリボザイム部分であることが証明されている。すなわち、タンパク質からできた酵素にその役割が移行したアミノアシルtRNA合成と異なり、現存の生物でも、実際にこのペプチド転移反応はリリボザイムが行っている。
図2に、現存の生物に見られるリボゾーム、tRNA、アミノ酸、ポリペプチドの関係を示したが、mRNAの塩基配列に対応したアンチコドンを持つtRNA同士が集まり、それぞれが担っているアミノ酸が重合する。この時、tRNA が集まる場所をリボゾームが提供していることがわかる。すなわち、図1で想定したマトリックスの役割をmRNAとリボゾームが演じている。これに加えて、リボゾーム内のリボザイムは、tRNAが担っているアミノ酸を順々に結合させ、ポリペプチドを合成するペプチド転移酵素作用も持っている。このように、原始リボゾームも現存のリボゾームも、tRNAを濃縮するマトリックスとペプチド転移酵素の集まったユニットと見ることができる。勿論RNAワールドでは、環境に適合するRNAは複製により増幅される(遺伝の可能性)ことは言うまでもない。
図2 現存のリボゾームの模式図。説明は本文参照。(ウィキペディアより引用)。
こう見てくると、タンパク質を作るためのすべての基本過程、すなわちアミノアシル化tRNAの合成及び、ペプチド転移反応の両方をリボザイムが行っていたことは重要だ。生命誕生過程が、先行して複製能力(遺伝能力)を実現したRNAワールドに、タンパク質が後から加わるという順序で進んだことを強く示唆している。言い換えると、RNAの活性がペプチドの合成も支配していた時期から、性質の異なる2種類の有機分子が互いに作用し合う世界へ移行したことがわかる。
物質として全く異なる核酸とアミノ酸を対応させるための記号化が必要になるのはまさにこのときだ。
原始型から現代のtRNAへの構造変化は情報誕生過程のヒントを与えてくれる。すなわち、原始型tRNAではアンチコドンはアミノ酸との物理的結合に使われており、情報を解釈する機能はない。一方現代型tRNAでは、アミノ酸と対応するアンチコドンが存在するが、これはアミノ酸と結合しているループから離れて存在しており、直接アミノ酸とアンチコドンが作用し合うことはない。 考えてみると、現存の分子で、物質と記号化情報がこれほど上手に媒介できる構造を持つ分子はtRNA以外にないだろう。しかも、RNAワールドではRNAの塩基配列により生まれる機能であれば複製・増幅が可能だし(遺伝)、多様化と選択(進化)も可能だ。
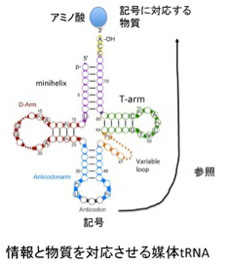
図3 情報と物質を対応させる媒体としてのtRNA)。tRNAの機能を知るため52話の図を再掲。
この段階で情報誕生の接点に存在する役者をリストすると、1)アンチコドン配列の持つ物理的性質の結果特定のアミノ酸に結合する原始型tRNA、2)ペプチド転移活性を持つリボザイム、3)異なるアミノアシルtRNAを集めるためのマトリックス、4)RNAを複製するリボザイム、4)RNAの安定性を高める、ランダムに形成されたポリペプチドと数種のアミノ酸だ。(図4)
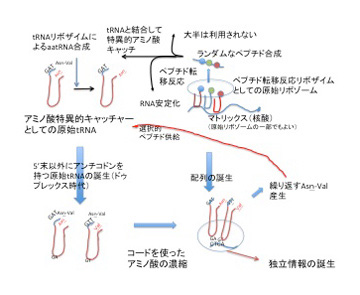
図4:遺伝と多様化が可能な情報の発生過程。説明は文中。
この組み合わせからどう情報(コドン)が発生したのか?
ここで仮に、清水のC4N仮説が正しいとしよう。この仮説では、特定のアミノ酸をキャッチするためにアンチコドンと、ペプチドが相互作用をしている。この時必要とされるペプチドはアスパラギン酸とバリンが結合したジペプチドだ。現代のコドンで表現するとGA(T/C)GT(T/C/A)の配列になる。
2種類のアミノ酸の組み合わせなら、ランダムでも合成するのは簡単で、生命誕生以前にも十分Asn-Valは存在したのではと思うが、 当時10種類前後のアミノ酸存在したと仮定してAsn-Valの組み合わせがランダムに合成される確率は、全ジペプチドのたかだか1%程度だけしかない。このペプチドを確実に十分な量作るためには、記号化されたコドンに従って、2種類のaatRNAを並べ重合させる必要がある。
ではどうすればこれが実現するか?
まずtRNAのアミノ酸と結合する場所(アンチコドンの物理化学的性質が利用されている場所)以外の部位にアスパラギン酸とバリンに対応するアンチコドンを持つ2種類のtRNAが必要だ。これに加えて、マトリックスとなるなんらかのRNA(一番理想的なのはマトリックス+ペプチド転移酵素として働く原始型rRNA自体)に6merのコドンが発生する必要がある。もちろんコドン/アンチコドンの形成は偶然の産物でなければならない。
6merだとこの配列が出来る確率がかなり落ちるので(1Kbに一回)、ここでは計算を簡単にするため、生命以前の原始の世界ではトリプレットコドンではなく、ダブレットコドンだったとしておこう。現在のコドンの冗長性からこう考える人も多い。この場合、tRNA側のアンチコドンはCTがアスパラギン酸、CAがバリンのコドンになる。
アミノ酸キャッチャーとして機能するtRNA5’末のアンチコドンや3’末のNCCA配列は、原始tRNAとしてRNAワールドでは安定に複製・遺伝が行われているとする。このtRNAのどこかにアスパラギンに対応する記号配列が現れる確率は、4×4(16mer)に一回になる。tRNAの大きさを50merと考えると、tRNAのどこかにこのアンチコドンが3箇所は存在することになる。
一方コドンを提供するマトリックスの方だが、ランダムにCTCAが合成される確率は4x4x4x4=256だ。もしマトリックスを提供していると想定している原始rRNAをやはり50merぐらいだと考えると、リボゾームの5個に1個はこの配列を持つことになる。
もちろんリボゾームとは別にRNA鎖を想定してもいい。リボゾームを考えたのは、すでにRNAワールドで複製が行われているRNAである必要があるためで、それが分解して出来た機能を持たない短いRNAでも、それが複製されるなら問題はない。実際、独立したRNAがリボゾームと結合してマトリックスを提供する方が、現在のリボゾームとmRNAの関係に近い。
いずれにせよ、RNAワールドで、ここでリストしたような変化が積み重なることは十分あり得るのではないだろうか。この意図せぬランダムな変化が重なり、2種類のaatRNAがこのCTCA配列を持ったマトリックス上で並んだら、全くランダムだった時の100倍の効率でAsn-Valを合成することが出来る。生産すべきペプチド産物を記号化して指示する独立した情報がここに誕生した。この過程を語る時、どうしても目的論的に語ってしまうが、実際には全てが偶然の過程として起こったことを強調しておく。
一旦この関係が成立すれば、ダーウィンが「もともと生命は、いろいろな力が、一握りの、ひょっとしたらたった一つの原型に吹き込まれて始まり」(種の起源 最終センテンス)と述べた、進化し続けることのできる情報が物質世界に生まれたことになる。この時点ではまだLUCAが誕生したわけではない。最初に吹き込まれたのは、情報だ。情報が誕生するということは、DNAやRNAが誕生することとは別の話で、物質でないコードが、特定の物質と参照関係を確立したということだ。これが達成されるとあとは情報自体が多様化し、さらに新しい大きなペプチド鎖を作ることができる。また、一種類だけでなく、2種類でも、3種類でもアミノ酸配列の決まったペプチドを増やすこともできる。要するにこれ以降は、ダーウィンが進化論で述べたのと同じだ。
最初情報に基づいて作られる特定の配列を持った短いペプチドの重要な機能は、例えば清水の仮説では原始tRNAにアミノ酸キャッチャーとしての機能を与えることであり、またペプチドのRNA保護作用を利用して、多くのRNAを安定化してRNAワールドの持続性と効率を促進することだっただろう。
しかし、できてしまえば多様化する情報断片の威力はすさまじい。こんな小さな情報でも、物理世界に自発的に誕生させることは難しいが、それでもなんとか十分説明が可能であることがわかっていただいただろうか?
今回は私がなんとかひねり出したシナリオを述べた。私でも捻り出せるぐらいだ。他にも十分可能性のあるシナリオが提出されている。次回は他のシナリオについて説明し、物理世界に情報が誕生することがそれほど荒唐無稽なことでないことを再確認したいと思っている。
[ 西川 伸一 ]
ゲノム誕生に関するRoot-Bernsteinの考え
2016年5月2日
ゲノムや生命の誕生について、これまで自分勝手な作り話に基づいて話を進めてきたので、「信用ならん!」と不快に思った読者も多いと思う。一つの仮説を皆さんに押し付ける意図は全くない。実際、38億年前に何が起こったのかを特定することはほぼ不可能だろう。期待したのは、多くの読者が、無生物から生物を発生させることが可能だと思ってくれることだ。
この文章を書き始めた時、無機分子から有機分子、そしてゲノムや生命の誕生など、考える糸口を見つけることすら難しいだろうと思っていた。ところが書き始めて1年もすると、生命誕生について合理的なシナリオを考えること自体はそんなに難しいわけではないと思うようになってきた。そして今は、21世紀にここで述べた様な幾つかのシナリオが必ずだれかによって実験的に確かめられると確信している。
繰り返すが、ゲノム誕生についての個人的理解をくどくどと語ってきた最大の目的は、無機物からの生命の誕生が、多くのシナリオが考えられる、当たり前の科学になりつつあることを皆さんに理解してもらうためだった。
そこで今回は、Root-Bernsteinという研究者が最近の論文で提案しているシナリオを紹介することで、前回述べたRNAワールドからゲノム情報が誕生するまでの過程をもう一度整理したいと思っている。ほとんどの読者にRoot-Bernsteinという名前は馴染みがないだろうが、シナリオは面白く説得力が高い。
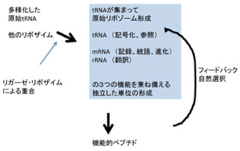
図1Root-Bernsteinの論文(Root-Bernstein & Root-Bernstein, J. Theo Biology 367: 130-158, 2015)に基づいて私が書き直したシナリオ。
まずこのシナリオについて、図1を使いながら簡単にまとめてみよう。このシナリオでは、原始リボゾームを、まだ生物ではないが、独自に、代謝、複製、情報による翻訳と進化の能力を獲得した独立した単位だと考えている。
すなわちリボゾームは、
の全てが備わった独立単位として考えられている。
エネルギーや物質代謝については、まだまだ独立しているとは言えないが、熱水噴出孔など自然の条件さえ整えば、Ottoの複製子のように複製、進化も十分起こりうる。しかも、複製時の情報としてだけでなく、アミノ酸に対応する記号化された情報も持っており、これにより様々なペプチド鎖を繰り返し合成し、また進化・多様化させることが可能になっている。例えると、適切な環境条件に寄生するウイルスが誕生したと言ってもいい。
これまで私の話に付き合っていただいた読者なら、Root-Bernsteinの仮説を荒唐無稽だと拒否することはないだろう。ただ、本当に生命誕生の途中段階で、ここに描いたような多様な機能を同時に表現したリボゾームが存在したことを証明できるかが問題だ。これについて、彼らは過去に存在した機能の痕跡は、現存のrRNAにも必ず見つかると考え、研究を続けている。
事実、1983年Blochらは現存のrRNAの配列内に、tRNAの配列の一部が頻回に現れることに気がつき、rRNAとtRNAは同じRNA分子から進化した可能性を示唆した(Bloch et al, J Mol Evol , 19:430-428, 1983)。この発見はその後他のグループでも確認されているが、Root-Bernsteinは彼らのシナリオを提案するにあたって、まずこの可能性を詳しく再検討している(Root-Bernstein and Root-Bernstein, J. Theretical Biology 367:130-158, 2015)。
図2は彼らの論文の図2、図3から転載したものだが、現存の大腸菌のrRNAにも、ほぼ全てのアミノ酸をカバーするtRNAの配列の一部が散らばっていることがわかる。
図2:現存の大腸菌の16S,23S rRNA配列の中に発見された、20種類のtRNA配列。(上記論文より転載)
さらに、こうして大腸菌rRNA内で特定したRNA配列が、tRNAの機能を持つかどうか立体構造を計算してみると、図3に示すように、現存のtRNAによく似た構造を取れることがわかる。
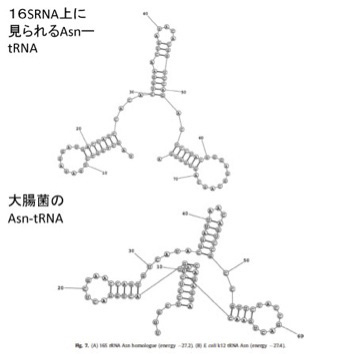
図3 rRNA内から抽出したAsn-tRNA(アスパラギン酸tRNA)の相同配列の理論的立体構造。実際のAsn-tRNAとよく似ていることがわかる。
このように、現存の20種類全てのtRNAに、配列上でも、構造上でも相同な配列がrRNAサブユニットの中に発見できるなら、tRNAとrRNAが同じ先祖RNAから進化したことは言うまでもなく、図1に示したようにrRNA自体がいくつかのtRNAが重合してできたものと考えた方が自然だ。
Root-Bernsteinらはこの点について明確には言明していないが、例えばdeFariasらは、まずアミノ酸との相互作用が可能になったtRNAが多様化し、次にtRNA同士が集まってrRNAのサブユニットを形成したというシナリオを提案している(deFarias et al, Life 6:15, 2016)。
図4はdeFariasらの論文から転載したものだが、tRNAは、その本来の機能であるアンチコドンも、また現在mRNAが担うコドンも両方備えており、それが重合することで、rRNAがmRNAとtRNAの両方の機能が備わった単位になるというシナリオを提案している。
さて、図1と図4をよく比べると、原始リボゾームをRNAワールド本来の代謝や複製に加え、転写翻訳のための全システムが備わった独立した単位として考える点では、図1で示したRoot-Bernsteinらの考えとほぼ同じと言っていいだろう。
図4 deFarias et alらの論文( Life 6:15, 2016)より転載した、原始リボゾーム形成過程の説明。
さてこのシナリオでは、原始リボゾームで最初作られていたタンパク質は、専らこのrRNAにコードされたtRNAやmRNA、そしてrRNAを安定化する作用を持っていたと考えられている。というよりは、安定化に関わるペプチドやタンパク質を作る情報を持つrRNAが自然選択されたと考えている。即ち、リボゾームが自然選択をうける一つの単位として完成していることがよくわかる。
では、本当にrRNAの中にタンパク質やペプチドをコードする配列は存在したのだろうか。例えば現存のrRNAには50種類以上のタンパク質が結合しているが、これらはrRNAにコードされているのではなく、rRNAをコードするゲノム部分とは異なる場所にコードされている。
実験の詳細は省くが、それぞれの配列の比較からRoot-Bernsteinらは、先に列挙した
の全てが、実際にrRNAの中に特定出来ることを示している。
このことは、最初rRNAの安定化に関わるタンパク質を合成していたリボゾームが、効率の悪いリボザイムに頼っていたRNA複製機能や、アミノアシル化tRNA合成機能をタンパク質に置き換える中心的役割を担ったと考えられる。
図5は、大腸菌の23S,16S,5SrRNA内にそれぞれ認められる現存のタンパク質をコードする配列の断片をプロットしたものだが、赤で示したリボゾーム結合タンパク質以外に、DNAやRNAポリメラーゼやtRNA合成酵素を含む多くの機能タンパク質遺伝子に対応する配列がrRNA内にこれほど認められるのは驚きだ。
図5:様々なリーディングフレームにおけるtRNA及びタンパク質の位置を示したマップ。
これらの結果を基礎に、Root-Bernsteinたちは、地球上のタンパク質の全ては、このrRNA内にコードされていた遺伝子に由来するのではと提案している。
以上、今回紹介したRoot-BernsteinやdeFariasらのシナリオをもう一度まとめると次のようになる。
ただ、rRNAはいくら脂肪膜で囲まれていたと言っても、生物ではない。次回は、このrRNAからLUCAへの道を考えてみたい。
[ 西川 伸一 ]
原始リボゾームからゲノムの誕生
2016年5月16日
前回紹介したRoot-Bernsteinの原始リボゾームの仮説は面白いだけでなく、生命誕生過程の構想に重要な示唆を与えてくれる。そこで今回はまずこの説を中心にしてこれまでLUCA誕生について議論してきたことをおさらいしておこう(図1)。

図1:LUCA誕生へのシナリオ。
説明は本文
先ず、熱水噴出孔でエネルギーと有機物の生成システムが形成され、ランダムではあっても様々な長さのRNAが持続的に供給されるようになる(図1 step1)。RNA自身は4塩基の組み合わせからできているが、多様な立体構造を取ることができるため、その構造の持つ触媒活性を通して、ランダムで多様化する有機合成反応を一定の方向に制約する可能性を持っている。例えば、一定の構造を持つRNA鎖が高い頻度で合成されるようになる可能性だ。
さらに、RNA同士を結合させるリガーゼ活性、塩基配列に基づいて複製するポリメラーゼ活性を持つリボザイムが生まれると、特定のセットのRNAを増幅・維持するRNAワールドが誕生できる(図1Step2)。RNAワールドの重要性は、何よりも塩基配列を鋳型として複製することで、同じ構造を増幅できる点、及び個別のRNA鎖が持つ触媒活性を媒介にして、相互に作用し合う分子群をひとつの単位として統合できることだ。
周りの条件にもよるが、RNAには安定性の問題がある。おそらくこの解決としてアミノ酸やペプチドとRNAの相互関係が始まったのだろう。ペプチドと言っても、当時自然に合成できたアミノ酸がランダムに結合したペプチドだったと考えられるが、それでも十分なペプチドを作るためには、環境中に低い濃度で存在しているアミノ酸を捕捉して濃縮する働きを持つ原始tRNAが必要になった。比較的安定なtRNA様構造を持ち、ペプチド合成の効率を上げてRNAワールドを安定化させるという利点を持つ原始tRNAが一旦RNAワールドに誕生すると、選択的に複製されることで自然に濃度を高めるだろう (図1step3)。そのうち、アミノアシルtRNA合成能力獲得してより安定にペプチドを作る独自の進化を遂げたと考えられる。もちろんペプチドを合成するためにはtRNA進化と並行して、アミノ酸転移活性を持つリボザイムもRNAワールドでは選択的に増殖したはずだ。
こうしてレパートリーを増やしてきたtRNAやアミノ酸転移活性を持つリボザイムが原始リボゾームへと統合される契機については想像でしかないが、リボゾームとしての機能発揮に必要なRNAを別々に複製するより、一つの単位として複製した方が都合のいいことは十分理解できる(図1step4)。
このような契機から、tRNAやリボザイムRNAが少ない数のRNA鎖に統合されて誕生したリボゾームは、エネルギーや物質代謝に関しては熱水噴出孔の環境に依存する寄生体と呼べるが、1)自己複製能、2)ペプチド合成能、3)そして翻訳のための鋳型としてのmRNA機能を持つ、「自己の誕生」とも言える一つの単位を形成する。
同じ自己複製能を持つOttoの複製子と比べると、自らを鋳型として複製できる点、ペプチドを合成する「翻訳機能」を持っている点で、はるかに高度で独立性の高い複製子がリボゾームとして誕生したと考えることができる。
以上が、deFariasやRoot-Bernsteinが考える原始リボゾームを中心とした大まかなシナリオだが、LUCAへの過程で重要な次の一歩は、情報の機能からの独立だと私は思っている(図1step5)。
RNAワールドでは、RNAが情報を担う媒体としての機能と、リボザイムとしての機能という二役を演じていることが重要だが、情報の独立とは、この一人二役が解消され、情報を担う媒体としてのRNA鎖と、様々なリボザイム機能を担うRNA鎖が完全に分離することを指す。事実現存の全ての生物のゲノム情報はDNAを媒体として、原則として情報以外の機能を併せ持つことはない。
情報が独立することは、ダーウィン進化にとって極めて重要な要件だ。例えば、DNAは情報の媒体以外の機能を持たない。このおかげで、細胞や個体という全体の中の一部であっても、全体からの制約を受けずに情報自体を比較的自由に変化できる。もちろん、情報が変化することで全体の維持が不可能になると、多様化した情報も消失する。とはいえ、情報の変化自体は全体の制約をほとんど受けないことが重要だ。
一方、現存のリボゾームの構造を見てみよう。図2はWikipediaから借りてきた50SリボゾームRNAの構造だが、RNAが折りたたまれ、一つの構造を形成しているのがわかる。
図2 50Sリボゾーム(Wikipediaより) 青で示されているのがリボゾームタンパク質で、残りはRNA.
この一部がmRNAの機能を併せ持つ場合、この部分は50SRNAの機能や構造の維持に必要とされ、この部分で起こる変異は即座にリボゾームの機能喪失につながる。即ち、情報としての自由な変化は最初から制約されてしまう。
このためダーウィン進化で見られたようなほとんど無限と言える情報の多様化がRNAワールドで可能になるためには、制約の源となる様々な機能から解放され情報に特化したRNA鎖が発生することが必要になる。そして最後に、RNAワールドでの配列情報を全てDNAに置き換えて、全体を情報に特化させる転換が必要になる。
この情報の独立が原始リボゾームでどのように起こったかについての私の妄想を述べよう。まずrRNA配列のどこかに、リボゾーム維持に全く影響しない余分な配列が生まれる(図3)。この部分はリボゾームとしての機能を持たないが、rRNAと一体化しており、複製もリボゾームの一部として行われる。このはみ出したRNA配列が、tRNAのアンチコドン部位が結合するコドンを提供するmRNAとして働きだすと、リボゾームの機能から独立した情報としてダーウィン進化が可能になり、多様な新しいペプチドが作られるようになる。すなわち、リボゾームの機能にとらわれず、様々なペプチドを合成して、自由に試してみることができるリボゾームが誕生する。
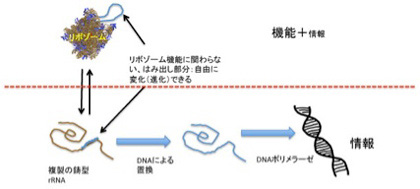
図3 情報媒体としてだけはたらく核酸の誕生。
しかし、おそらくこのはみ出し部分だけが情報媒体として独立することはないだろう。というのも、このはみ出し部分がmRNAとして機能するためには、リボゾーム全体の存在が必須だ。このため、このはみ出し部分は、rRNA以外のペプチドを作るための進化する情報として存在しえても、rRNAから独立することはできない。
したがって、完全に独立した核酸を媒体とする情報が誕生できたとすると、この情報はrRNA(その内部のtRNAも含む)をはみ出し部分と合わせてコードする必要がある。例えば、rRNA配列全体を全てDNAに置き換えることができれば(すなわち逆転写が起これば)、一挙に情報に特化した核酸媒体を誕生させることができる(図3)。しかし、逆転写酵素、あるいは逆転写リボザイムが都合よく突然現れて、情報を独立の問題を一挙に解決すると考えるのは虫が良すぎる。
これまで触れてこなかったが、実を言うとRNAワールドでも、複製を維持するためには、機能のない相補的RNA鎖が作られ、それを鋳型として機能的RNA鎖を合成する必要がある(図3)。従って、機能の制約を受けない情報に特化したRNA鎖がRNA ワールドでは常に合成されていたことになる。ただ、機能を発揮するための構造化ができない RNAは不安定で、おそらく情報として長続きできなかったはずだ。
ここでRNAポリメラーゼ活性を持つリボザイム自体のヌクレオチド特異性が高くないため、複製時にDNAが取り込まれることもあったと考えればどうだろう。リボゾームとして機能するRNA鎖は、多くのDNAが混じると機能低下につながるだろうが、それでもかなりの機能を維持できたと考えられる。実際、RNAとDNAが混合したアプタマーの機能が現在研究されており、これもリボザイムより安定であるという性質を期待しての研究だ。
一方、複製の鋳型としての機能は、DNAが混じるほど安定化し寿命が伸びる結果、DNAで置き換わった程度が高い鋳型の濃度が上昇し、最終的に完全にDNAを媒体とするゲノムが誕生したと考えられる。すでに述べたように、脂肪酸だけでもDNA複製が自然に起こることがある。一旦情報媒体に特化したDNA鎖が生まれれば、機能を持つrRNAも、多様化するために生まれたmRNA部分も、全てが一つの情報としてまとまる。ゲノムの誕生だ。
ゲノムが独立することで、この複製単位はリボゾームの範囲から拡大し始める。次回はこの自己の範囲の拡大について見ていく。
[ 西川 伸一 ]
記号、イコン、インデックス、シンボルと情報
2016年6月1日
しつこいようだが、今回もこれまでの話を繰り返してまとめ直すことにした。もともと生物がどう誕生したかについては、本当の答えを導き出すことは難しい。従って、理論的に説明可能かどうか、何度もなんども考えて納得できるシナリオを書いていくしかない。また説明するときも、これまで議論した過程を表現し直して、なんとか読者の皆さんにも理解していただきたいと思っている。もちろんまったく同じ説明を繰り返すつもりはない。様々な視点から同じ過程を説明し直すことが重要だ。
今回はLUCA誕生へのプロセスを、情報と記号論の観点から見直してみようと思っている。すなわちLUCA誕生の過程でどんな情報がいつ誕生したのかまとめてみたいと思っている。この作業は、ゲノムの誕生を説明するのが目的だが、生物進化とともに新たに現れてきたまったく異なる情報、例えば人間の言語の誕生も視野に入れている。
テレオダイナミックス系の誕生
これまで生命誕生へのスタートラインを、エネルギーと有機物が持続的に合成される熱水噴出孔に生まれた多種類の散逸系の誕生と考えて来た。この過程で有機高分子が発生すると、様々な分子間の相互作用が起こりやすくなり、本来は無秩序な物質やエネルギーの流れに一定の秩序がもたらされる。ただこの分子間の相互作用による秩序や規則性の誕生には、どのような意味でも情報と呼べるものは存在しない。Ottoの複製子で見たように、偶然発生した有機分子が、エネルギーや分子の合成過程を制約して、自己複製まで可能になることもある。しかし、これは分子と分子の直接の相互作用による制約であり、情報とは呼べない。渦を持続的に発生させる流れの中に置かれた石と同じと考えればいいだろう。ただ、この制約は偶然の産物で、物理法則による制約とはまったく異なる。熱水噴出孔で起こっていることをもう一度まとめると、比較的安定な様々な制約要因と、それに制約された「傾向」が生まれ、この傾向が蓄積することで一定の秩序を持った系が生まれるといえるだろう。傾向は情報ではない。これをDeaconはテレオダイナミックス系と呼んだ(図1)。

図1:テレオダイナミックス系の誕生(2月15日の記事とほぼ同じ)
イコン的記号の誕生
この制約された有機物合成システムからヌクレオチドが合成され、それが重合してポリヌクレオチドが生まれる。RNAポリヌクレオチドの中には様々な酵素活性を持つ物が存在し、リボザイムと呼ばれており、現存の生物の中でも実際にリボザイムが働いている。このリボザイム活性は、反応の制約要因としてはより高度の制約活性を持ち、様々な分子の合成過程に秩序をもたらす。それぞれのリボザイムは塩基が並ぶことでできている。この塩基配列は記号と呼んでいい。しかし、記号だから情報というわけにはいかない。RNAが結局限られた数の異なる塩基の並びでできているため、リボザイムがこの塩基配列に支持されてできているように見えてしまうが、合成自身はランダムに起こっており、何かを指示する情報は存在しない。
このように、塩基配列という記号が直接分子の物理的構造を決める場合、記号論的にいうと、イコン的記号性が発生したと言える。
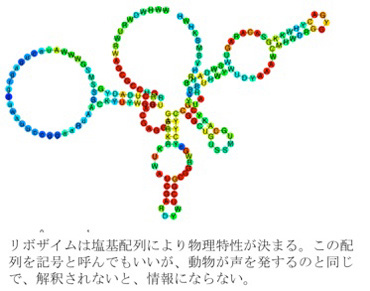
図2イコン的記号の誕生。 リボザイムは塩基配列により物理特性が決まることから、配列(記号)が分子構造や機能と対応する点でイコン的記号の誕生と言える。
情報とインデックス記号の誕生
ところが、リボザイム活性を持つRNAポリヌクレオチドの集団の中に、特定のRNA配列を認識してRNA鎖を結合させるリガーゼ活性や、更には塩基配列に従って複製するRNAポリメラーゼ機能を持ったリボザイムが生まれると、RNA鎖の塩基配列が突如反応を指示する鋳型としての機能を持ち、鋳型に従って同じ分子や構造を正確に再生産することが可能になる。具体的には、同じ分子構造を、塩基配列(記号)を鋳型(情報)として繰り返し合成できるという点で、記号がついに情報になったと言える。
このシナリオからわかるのは、リボザイムを構成する塩基配列が分子の構造を決めているとしても、この違いを解釈する仕組み(すなわちRNA ポリメラーゼ)が誕生するまではイコン的記号で、情報性は持たないことだ。言葉を変えると、塩基配列を鋳型として使って、その鋳型に合わせて合成を行う酵素やリボザイムが誕生すると(解釈者の誕生)、塩基配列(記号)に情報としての機能が生まれる。 ではなぜこの塩基配列を情報と呼べるかというと、この鋳型はRNAだけでなく、DNAでも表現できる。しかし同じ配列がDNAを媒体として表現されると、RNAで表現できる構造とはまったく異なる構造が生まれる。一方、鋳型としての機能はRNAでもDNAでも同じだ。この物理的媒体を選ばない点で、複製される塩基配列は情報と呼んでいい。
まとめると情報の誕生とは、塩基配列という記号の誕生ではない。記号が解釈されて初めて情報になる。この意味で、地球上で最初に情報が誕生した時とは、RNAポリメラーゼ活性を持ったリボザイムが誕生した時と言えるかもしれない。こうして誕生したRNAの構造と塩基配列の関係は、塩基配列(記号)が、リボザイムの持つ構造や機能のインデクスになっている点で、記号論でいうインデックス記号と言ってもいいのではと思っている(パースなどのインデックス記号の定義とは異なるが)。
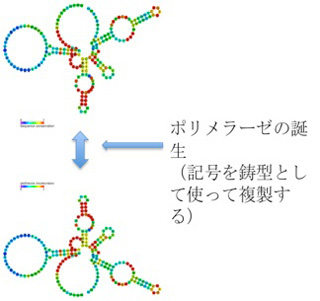
図3:インデックス記号と情報の誕生
ただこのインデックス記号は、まだ化学的に無関係な物質同士が対応関係を持つ、すなわち本来のRNAとはまったく無関係のアミノ酸がRNA配列と対応する段階、すなわちシンボル的記号には到達していない。(シンボル的関係の代表は言語で、サクラという音の並びと実際の桜には全く実体的な対応関係はない。)
シンボル的記号の誕生
ここで現存の生物が全て持っているDNA-RNA-タンパク質というセントラルドグマを考えてみよう。DNAの塩基配列(記号)は、RNAの塩基配列に転写される。これは、RNAワールドでの複製機能と特に変わることはない。しかし、転写されたRNAを鋳型としてタンパク質が作られる過程は、DNAの塩基配列とtRNAと結合するアミノ酸配列がもともと何の関係もないという点で、塩基配列がシンボル的記号に転換している。
この塩基配列という記号がアミノ酸と対応してシンボル的記号に転換するためには、まず最初にtRNAの誕生が必須だ。ここでは原始tRNAが、RNAワールドを安定化するための短いペプチドを安定的に供給するため、アミノ酸を捕捉して濃縮する目的で生まれたことにしている。
すでに述べた清水らの説に従えば、原始tRNAが持つアンチコドン配列と、たまたま存在したAsn-Valペプチドが相互作用すると、特定のアミノ酸が捕捉される。ただここで使われるアンチコドン配列(記号)は、決して情報として使われているのではない。ペプチドとともに、アミノ酸を捕捉するポケット構造の部分として使われており、アンチコドンと呼ぶのは、現在の生物と比べて言っているだけだ。したがって、このアンチコドン配列によるアミノ酸把握で配列はイコン的記号として使われているが、情報としての関係はない。
とはいえ、このようなアミノ酸とアンチコドンとのイコン的関係が成立することで、原始tRNAに捕捉されたアミノ酸を濃縮することができ、情報でなくともペプチド合成の効率をあげることができる。この時、原始tRNAが集まるためのマトリックスとして、RNAやDNA鎖が働いていたことは十分考えられる。だとすると塩基配列という記号が、RNAとは全く物理的関係のないアミノ酸と対応したシンボル的記号へ転換するためにはあと一息だ。
原始tRNAがペプチド合成のためのアミノ酸捕捉分子として機能するためには、複数のアミノ酸とイコン的関係を持つ原始tRNA、アミノ酸をtRNAに結合するアミノアシル化活性を持ったリボザイム、集まったアミノアシルtRNAに結合したアミノ酸同士を結合させる、アミノ酸転移酵素活性を持つリボザイムは最低限存在していなければならない。特定のアミノ酸が結合したペプチドが持つRNA保護作用や、一部のペプチドによるリボザイムの機能の効率化などが強い選択圧として働き、このようなリボザイムが化学進化したことは十分考えられる。
この前提が満たされた時、核酸配列という記号がアミノ酸配列と対応するシンボル的記号へ転換する条件が整う。その後のプロセスを順を追って考えてみよう(図4、5)。

図4:本文と同じことを箇条書きにしている。これ以外にも多くの合理的可能性がある。
まず最初ただのマトリックスとして使われていたRNA(ここでは便宜的にrRNAとしておく)とtRNAの関係が、コドンとアンチコドンが持つより強い結合を基盤にした関係へと移行する必要がある。rRNAやtRNAにアミノ酸をコードするコドンを見つけることは難しくないが、実際にはその配列がどこにあってもいいというわけにはいかない。rRNA上の配列をコードとして集まったtRNAに結合したアミノ酸同士が結合するための十分近い距離に保つ必要がある。
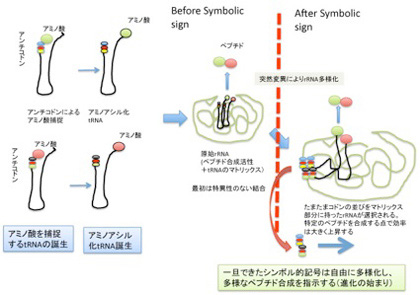
図5 原始tRNA誕生からシンボル誕生まで。説明は本文。
例えば、まずアミノ酸転移反応を制御するrRNAと、tRNAの構造がともに進化して、記号対応関係はないがアミノ酸が濃縮できるマトリックス構造が生まれた後、この結合をさらに強い結合にするために、rRNAの一部がコドン配列を特定の場所に持つように進化すると考えてみよう。
この時、清水らの説のように、アミノ酸をキャッチして3’端にエステル結合させる過程で、tRNA5’末のアンチコドン配列が物理的(イコン的)記号の働きができるとかなり話が早い。すなわち、アミノ酸補足後アミノアシル反応が終わり、3’末にアミノ酸が結合した後、フリーになったアンチコドン部分がマトリックス側に現れたコドンと結合して、tRNAの結合を強め、アミノ酸転移反応の効率をあげると考えれば、選択されるのはrRNA側の配列だけになり、考えやすくなる。すなわち、tRNAのアンチコドンは最初からイコン的記号として存在し、rRNA側のコドンが選択された時に初めてシンボル的記号に移行すると考えるのは、そう難しくない。
このシナリオの問題点は、マトリックス側に偶然現れるコドン配列の長さに限界があることだ。実際、2−3アミノ酸分が関の山だろう。とすると、必要なペプチドごとにそれに対応するコドン配列を持ったrRNAが必要になってしまう。
これについての一つの答えは、まず「簡単なものから始めて見る」ではないかと思っている。すなわち、実際多様なrRNAが存在する中で、RNAワールドにとって最も必要なペプチドに対応するコドンを持ったrRNAが選択的に増殖したと考えればいい。短くてもシンボル的記号としてのコドン部分が選択されたrRNA上に生まれれば、この部分は増大し多様化できる。
例えば最初単純な言葉が生まれ、そこから複雑な言語が発達するのと同じだ(この点については後々考察する)。簡単な記号を元にシンボル的関係を成立させることがまず重要だ。
あとはこのコドンを持ったマトリックスを独立させ、DNA化することで生命の情報システムは完成する。この過程についての一つの可能性はすでに前回述べた。
以上のシナリオをもう一度記号論的にまとめると、
今回は生命誕生について、生命情報成立をより強調して説明した。次回は原始リボゾームからLUCAへの過程について考えるが、この記号論は、今後様々な生命情報を考えていく上で共通の基準をあたえてくれる。
[ 西川 伸一 ]
進化の始まりと目的の実現
2016年6月15日
前回のまとめは理解していただけただろうか?
特に、核酸配列がイコン(類似)記号、インデックス(指標)記号、シンボル(象徴)記号へと発展した点についての説明は理解して欲しいと思っている。

図1チャールズ・サンダーズ・パース
我が国では語られることが少ないが、科学及び哲学の広い範囲にわたり現代のアメリカ思想の源を作った。その記号論はこれまで言語学の人によく引用されている。
この稿で、核酸配列をイコン、インデックス、シンボル記号に分類したのは、言うまでもなくアメリカの巨人チャールズ・アレクサンダー・パースの考えをそのまま拝借している。パース(図1)はプラグマティズムの創始者として紹介されることが多いが、一つの枠で語れない偉大な巨人で、その記号論は言語や核酸配列などの生命情報を整理する上で欠かす事が出来ない(ぜひ読んで欲しい)。
RNAに、4種類の塩基から出来ているという記号性と、それ自身で酵素活性を持つ2面性が存在することが、複製可能な有機システム=RNAワールドを可能にしたが、このとき最初からRNAが情報として働いていたと思うと大きな間違いを犯す。
配列が情報としてRNAポリメラーゼに解釈されようになるまでは、配列=構造という単純な物理関係があるだけで、この時の配列は記号ではあっても情報でない。このことから、これをイコン記号とした(図2)。
ここにRNAポリメラーゼが誕生すると、塩基配列を解釈する過程が始まり、塩基配列がインデックス記号に変化する。最後に、アミノ酸配列と核酸配列という全く無関係な物質同士がtRNA+rRNAの誕生で対応するようになると、塩基配列はシンボル記号になる。重要なのは、イコン記号及びインデックス記号の段階では、記号(塩基配列)とそれに対応する機能分子との間には、物理的法則に基づく対応関係があるだけだ。ところが、核酸配列がシンボルの記号になると、核酸配列とは全く関連性のないアミノ酸配列に記号が対応する。このことは、物理化学とは異なるルールがこの記号に導入されたことになる(図2)。

図2 パースの記号論を生命誕生に当てはめてみる。
勿論非物理的対応関係を媒介するのはtRNAとrRNAという現実の分子だが、核酸配列がシンボル記号としてアミノ酸配列に対応するようになるためには、これまで述べたような固有の歴史を通してそれぞれの関係性が生まれる必要があり、この歴史性は物理化学的法則にはない。このように、シンボル記号誕生こそが、物理科学法則に支配される地球に、新しい情報という因果性が誕生したことを意味する。そして、地球上にダーウィン進化が始まる。このダーウィン進化の始まりについて見ていこう。
核酸配列がイコン記号として働く段階では、同じ記号(=物質)が生まれるかどうかは偶然に支配されており、できる分子は無秩序に多様で、この段階の有機物質やエネルギー合成は無秩序なカオス状態と言っていい。ここに、核酸配列が複製のためのインデックス記号として働く情報化が起こると、分子の多様性を減らし、秩序をもたらす方向に進む。このように、もともと情報とは系に秩序をもたらす方向に働く。ただ、秩序を生み出す複製過程においても、一定の制約の中での複製エラーによる変異が核酸配列に起こる。この秩序という制約の中での多様化こそがダーウィン進化の原動力になる。
このように、生命とは呼べないが、原始リボゾームのような複製する原始ゲノムが誕生した時代を考えてみよう。RNAで代謝の多くをまかなうことができるとはいえ、有機物生産システム、エネルギー生産システムなど多くの生命に必須の条件はRNAのみで達成できない。従って、RNAワールドをコードする原始ゲノムは、環境の中に寄生したRNAウイルスのようなものとして考えればいい(しかし、私たち人間を含む現在の生命も結局環境に寄生しているが)。お分かりのように、私は生物の始原はゲノムから始まると考えている。
もともと生命をDNAから考える有名な仮説がリチャードドーキンスの「利己的遺伝子」仮説だ(図3)。現存の生物の進化について利己的遺伝子仮説が当てはまるかどうか議論がつきないようだが、LUCA誕生直前の状態を考えれば、妥当な仮説だと私には思える。

図3 リチャードドーキンスと著書「利己的遺伝子」
即ちLUCA誕生前の状態を、熱水噴出孔の特殊な環境に寄生しているRNA/DNA寄生体(利己的遺伝子)が、この環境から自立する過程として捉えればいい。未だ生物は誕生していないが、将来のゲノムの核になる原始ゲノムは既に存在している。この原始ゲノムが自立するためには、環境を形成している様々な分子を自分の中に取り込み同化する必要がある(図4)。
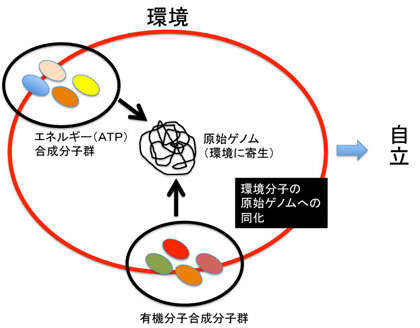
図4:環境に寄生する原始ゲノムの自立
環境を形成する分子がRNAやDNAでできている場合は、その配列を融合により同化すればいい。そうすることで、環境分子は複製により安定して調達されるようになり、自然と原始ゲノムのゲノムサイズが上昇する(図5)。
環境分子が核酸でなかった場合、原始ゲノムを多様化して新しいリボザイムを合成し、環境分子の機能と一番フィットした原始ゲノムを選択することで同化が可能になる。
もちろんこの様式で環境分子を同化してもいいが、RNAワールドには限界がある。RNAワールドでの核酸配列はインデックス記号であり、記号がそのまま物理的性質と直結する。このため、核酸レベルの構造が制約となり、多くの環境分子に対応する機能分子に対応する記号を合成することは制限される。
RNAだけで環境分子を同化していくことには他の問題もある。ゲノムサイズが大きくなると、ポリメラーゼ・リボザイムと相補的塩基配列がペアリングする危険性が高まり、複製の効率が落ちると予想できる。また既に述べたように、RNAの安定性を高め、機能を補完するためにもペプチドが必要になる。従って、原理的にはRNAだけで環境の同化を進めることは可能だが、核酸配列が物理的に制限されることなく機能や物質と対応できるシンボル記号へと転換することで、更に自由な同化が可能になる(図5)。
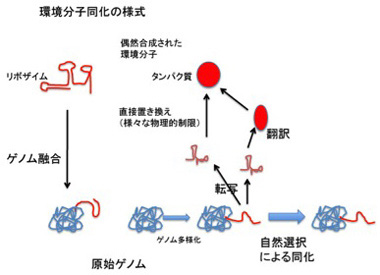
図5:環境分子同化の様式
この同化を繰り返せば、環境依存性が少しづつ低下していく。すなわち、より多様な環境で存在することが可能になる。これを繰り返してLUCAができると考えること自体は、私は、論理的にも無理はないように思っている。
面白いのは、RNA機能分子を原始ゲノムに融合する様式では、自然選択は全く必要ない点だ。一方、原始ゲノムを転写し、その産物で環境分子を置き換えられるかどうかを試す様式では、最も環境分子に近い分子を合成できる原始ゲノムが自然選択される。これは、水平遺伝子伝搬とダーウィン進化の共同作用による進化のルーツのように私には思える。
最後に強調したいのは、LUCA以前の、環境分子同化過程でのダーウィン進化では、原始ゲノムが同化すべき環境分子がすでに存在している点で、これは生命誕生後のダーウィン進化と決定的に異なっている。
ダーウィンの進化論は、目に見えない因果性、すなわち「目的」に駆動されているように見える生物進化が、多様性の獲得と自然選択のダイナミズムで説明できることを示し、最初に目的が存在することを拒否している。一方、原始ゲノム(利己的ゲノムと呼んでもいいかもしれない)が環境から自立しようと進化するとき、最初から同化すべき環境分子が目的として実在している(図4)。もちろん最終的にゲノムが同化された記号は、必ずしも同じ分子に対応する必要はなく、機能が維持できればいい。しかし、最初から手本として、目に見える分子が進化の方向を示す目的として存在していたことが、LUCA誕生前後のダーウィン進化の大きな違いであることは強調しておきたい。
あと生命誕生まで、自己という単位が、原始ゲノムから細胞へと拡大する過程が残っているだけだ。
[ 西川 伸一 ]
生きた細胞の再構成は可能か
2016年7月1日
これまで断片的にではあるが、生命誕生を考えるための様々な条件、あるいはシナリオについて解説してきた。多くの読者の方が、「条件さえ整えば無生物から生物が生まれる過程を構想することは全く不可能ではない」と考えてもらえる様になったのではと期待している。
このシナリオで最も重要なゴールは転写(ポリメラーゼ)機能+翻訳機能(tRNA+rRNA)を備えた原始ゲノムの誕生だ。もちろん原始ゲノムの誕生には、その維持と複製を支持する環境条件が必要だが、一旦誕生すると、この原始ゲノムはオーガナイザーとなって、環境を構成する分子を一つ一つ同化し、環境から独立しLUCAに至る。原始生命と呼べるのはLUCAが最初だが、最も重要なスタートラインは原始ゲノムの誕生と考えている。
40億年前の地球で起こった詳細をこのシナリオが説明できているとは到底思えないが、環境がまず形成され、それに寄生する原始ゲノムが誕生し、環境を自己に同化するという順番はおそらく間違っていないのではと思っている。
このシナリオでは、生命誕生に必要な分子は、もともと熱水噴出孔での「有機化学法則」に従って、個別に合成されるもので、各過程は独立に進み、組織化されてはいないと考える。従って、熱水噴出口の中のマイクロセルには、たまたま存在する分子ごとに多様な環境が形成されている。その中の一部の条件が、RNAワールド(イコン・インデックス記号的段階)から原始ゲノムへの進化の発展を支えることが出来たのだろう。これは全て熱力学と、非平衡熱力学が支配する過程と言っていい。
原始ゲノム誕生までの条件が地球上に揃う過程を頭の中で考えるのは簡単だが、これが実現するために必要な有機物の合成が維持される環境を、地質学的に十分長い時間維持することはそう簡単ではない。その意味で、地質学的時間一定の状態を維持できる熱水噴出孔の存在は欠かせない。この時、原始ゲノム誕生までに最低限いくつの分子が必要か決めることは、重要な課題だろう。というのも、タンパクや脂質の合成が互いに制約しあって一定の秩序を持って行われるとしても、この段階までの合成は原則ランダムで組織化されていない。必要最低限の数があまりに多いと、ランダムな化学合成ですべてを揃える確率は当然低下する。
ではいくつの分子が原始ゲノムの独立に必要だったのだろう?
実験室の環境で自律的生物を維持できる最小限の遺伝子数については、CJ Ventorのグループが精力的に研究を続けている。全部あわせても517個の遺伝子しか持たないマイコプラズマにトランスポゾンを感染させ、多くの遺伝子を同時にノックアウトする方法で、どの遺伝子が生存に必要かをしらみつぶしに探索した研究が1999年にScienceに発表された(Hutchinson III et al, Science 286:2165, 1999)。
たかだか265−350個の遺伝子で自律生命が維持できるというVenter達の結論には、無生物から生物への再構成過程を研究できるのではと私も興奮した。この数は、その後トランスポゾンの挿入された個々の遺伝子を丹念に検討し直す研究から、382個へと上方修正されている(Glass et al, PNAS 103:425, 2006)。それでも400弱の遺伝子があれば、実験室の環境で自律生命が可能であると理解すると、生物合成が手の届くところにきた様な錯覚に陥る。
しかし、生命誕生に必要な最低限の遺伝子数が400だとしても、これらが全て揃って地球上に生命が誕生できたようには思えない(計算根拠はない)。即ち非平衡熱力学過程から秩序がもたらされるとしても、億年単位の時間内に400全ての分子が地球の特定の場所に同時に出現する状況を考えるのは難しい。
幸い、熱水噴出孔の物理的条件の安定性については信頼できる。このため、原始ゲノムの存在した環境形成には、合成されたRNAやペプチドだけでなく、熱水噴出孔自体が提供する条件(例えば無機質の触媒作用や、プロトン勾配など)の果たした役割が大きいと思われる。従って、原始ゲノムが独立するまでには、環境を形成するRNAやペプチドの同化だけではなく、新しい機能を持ったRNAやペプチドを合成する情報を獲得することが必要だったと考えている。もし原始ゲノムがこの課題をクリアーできるだけの十分な能力を備えていると確信できれば、LUCA誕生は説明できたのと同じだ。
記号性を持ち、この記号を拡大・複雑化できるとともに複製能を持つ原始ゲノムは、生命ではなく、まだ環境に寄生しているが、多様化、自然選択、選択された個体(原始ゲノム)の複製を通したダーウィン進化を行う能力が備わっており、この過程を介して、環境の分子を同化し、また新しい分子を開発して環境から独立して生命誕生へと発展できる。すなわち、ダーウィン進化が可能な原始ゲノムが誕生した時点で、生命誕生のための基盤は全て整った。
ここで一つ思考実験を考えてみよう。生命最小単位マイコプラズマを集めて破砕し生きた細胞を消滅させる。こうして用意した、細胞内に存在する全ての分子が最適な濃度で維持されたマイコプラズマスープを、必要なら栄養やエネルギーを外部から適宜加えて維持できるとしよう。ゲノムの複製、転写、タンパク質の合成などが十分可能なスープだ。そこに合成された完全なマイコプラズマゲノムを投入すれば、もう一度生きたマイコプラズマが発生するだろうか?もし原始ゲノムがLUCAに発展するなら、マイコプラズマが発生しても不思議はない。
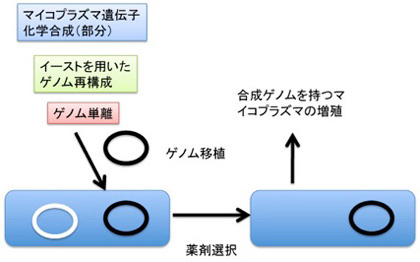
図1 CJ Venterの人工合成ゲノムを持つマイコプラズマ作成実験の概要
私の答えはノーだ。マイコプラズマの細胞質の条件を全て維持でき、その中でマイコプラズマゲノムが複製できたとしても、生物は生まれないだろう。なぜなら、この条件で、ゲノム自体がマイコプラズマの自己とは何かを決める事が出来ないからだ。一方、マイコプラズマのゲノムを外来のゲノムで置き換える実験は行われている。
先ずVenterのグループが行った、マイコプラズマのゲノムを置き換える実験を見てみよう。今年、Venterグループは完全に合成したゲノムを持ったマイコプラズマを作るのに成功して話題を呼んだが(図1:Hutchison III et al, Science 351:6280, 2016)、この時人工合成したゲノムを異なる系統のマイコプラズマに移植し、合成ゲノムを持つマイコプラズマを作成する実験を行っている。このマイコプラズマへのゲノム移植はどう行われたのか見てみよう。
詳細は省くが、合成ゲノムは先ず他の系統のマイコプラズマに移植されている。この時、移植した宿主マイコプラズマが持っていたゲノムを除去する操作は、この実験では行われていない。即ち、合成ゲノムを移植した段階で、宿主のゲノムと、移植した合成ゲノムの両方が共存する時期を経て、薬剤選択により宿主のゲノムが排除され、移植ゲノムが残る方法を用いている(図1)。実際にはこの方法は開発にはずいぶん苦労したようで、その難しさは2007年に発表された論文から窺い知ることが出来る(Lartigue et al, Science 317:632, 2016)。
ただここで注目したいのは、この苦労話ではなく、単離したゲノムは先ず生きているマイコプラズマに移植しないと、ゲノムとして働けないことだ。すなわち、ゲノムが細胞内の様々な過程を組織化するための情報として働くには、細胞が先に生きている(=組織化されている)状態を維持していることが必須である点だ。「卵が先か・鶏が先か?」の生命特有の問題がここでも現れる。
では、宿主のゲノムを除いても、合成ゲノムを移植することは可能だろうか。私は可能だと思っている。しかし、バクテリアから細胞質や細胞膜を傷つけずに残してゲノムだけを除去することは技術的に極めて難しく、このような挑戦が行われていないだけだ。
事実、細胞の操作が可能な十分な大きさを持つ脊椎動物の卵子では、核を取り除いた後、他の細胞から単離してきた核を注入し、ゲノムが完全に入れ替わったクローン動物を作ることが出来る(図2)。
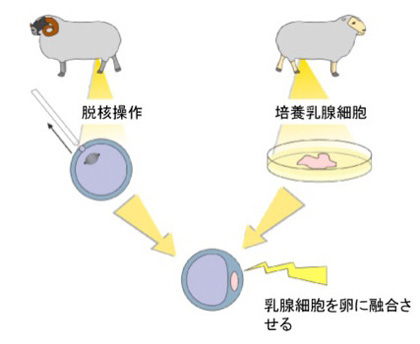
図2:クローン羊ドリーの作成。 まず卵子の核を除去した後、乳腺細胞を融合させている。
更に当時理研・発生再生科学総合研究センターの若山らは、細胞としては死んでいることが確認された脳細胞から核を取り出し、それを卵子に移植する事でクローン動物が作成できることを示し、ゲノム情報の維持には必ずしも細胞が生きている必要がないことを示している。従って、生きた細胞を宿主とすれば、宿主のゲノムの有無にかかわらず、ゲノムは働くことが出来る。
ではなぜ原始ゲノムは熱水噴出孔の環境で生命へと発展できるのに、マイコプラズマゲノムはマイコプラズマスープの中で生命へと発展できないのか? また、マイコプラズマスープの中で新たなマイコプラズマが出現しないのなら、熱水噴出孔の環境がどれほど完全でも、原始ゲノムからLUCAが発生することが、本当に出来るのだろうか?これを理解する鍵が自己性だ。
マイコプラズマスープにマイコプラズマゲノムを混合しても生命が発生しない理由を考えると、原始ゲノムと違って、マイコプラズマゲノムは進化の過程で情報に特化してしまい、既に生物の部分になってしまっているからだと言える。
すなわち、原始ゲノムはそれ自身で自立しており、自己性=全体性を持っている。一方、マイコプラズマゲノムはマイコプラズマスープの中で、複製し、タンパク質やRNAやタンパク質を合成出来たとしても、自己性=全体性は失ってしまっている。即ち、マイコプラズムの自己は、ゲノム、細胞膜、細胞質も含めた全体へと拡大している。単細胞生物の自己は、細胞膜で囲まれた内側にあるゲノムも含めた全てで、決してゲノムだけでない。このため、生きた細胞にゲノムを移植する方法で自己の範囲を明確に与えるという過程を踏まないと自己性は生まれない。細胞質スープの中でゲノムが自己組織化を通して細胞としての自己を自然発生させる可能性はあるにしても、その確率は低い。
少しわかりにくい議論になったが、進化により自己性の範囲が拡大することは、我々人間を含む高等多細胞動物を考えるともう少しわかりやすい。
例えば人間の自己は、決して身体の範囲内に制限されているわけではない。例えば私が今書いている原稿も、やはり私の一部だと言える。一方、意識がなくなった私の身体は、生きていたとしても人間としての自己性を失っている。すなわち、神経ネットワークというゲノムとは異なる情報を獲得することで、生物の自己の範囲は広がった(これについては、ゲノムの進化について考えるときもう一度考える)。原始ゲノム、LUCA、そして人間と、自己性が生物の全体を決め、進化と共に自己性の範囲を拡大させ環境を取り込んできたのが生物だ。
この自己性の拡大とは、環境の自己への同化の過程とみることができる。そしてこのことが最もはっきりわかるのが、原始ゲノムが環境を同化していく過程だ。要するに、情報性と自己性を持つ原始ゲノムが誕生すると、原則として無限の新しいタンパク質を作る能力が得られる。ただ、自己性の問題からわかるのは、完全な自己性を維持しているゲノムだけが、環境を統合した拡大自己を発展させることができる点だ。
一端ゲノムが自己の部分になってしまうと、もうゲノムが自己性を決める事は出来ず、従って生物は発生しない。ゲノムは今後は、自己のなかで、情報を担う部分として進化の駆動力として、多様化と複製に関わる。
どうだろう。生物が生まれる一つのシナリオは理解してもらえただろうか。もし疑問があれば、これからはそれに答えることも行いたいと思っているので、問題点を指摘してほしい。
ゲノムは誕生した。次回からはゲノムやゲノム以外の情報の進化について考えていく。
[ 西川 伸一 ]
ゲノムの進化と生物の進化
2016年7月15日
ゲノムの解剖学に始まり、ゲノムの生理学、そしてゲノムの発生学と、すでに27回にわたってゲノムに関する知識を独断で、しかもほとんどは自分の頭の整理のために書いてきた。特に後半17回はゲノムの発生学と称して、今私たちが知っているゲノムが、生物のいない地球でどう誕生したのか、思いつくまま書いてきたが、そのおかげで自分では無生物から生物が生まれる過程を頭の中で思い浮かべられるところまで整理ができたと思う。ただ、説明自体は体系的でなかったので、読者の方にはわかりにくかったのではと反省している。私の個人的な頭の整理に付き合っていただいた読者の皆さんにはまず感謝。
私自身は納得できたと満足していても、読者の多くは生命誕生の話がまだ完結していないという印象を持たれたかもしれない。事実、ゲノムの発生学についてのシナリオは原始ゲノムまでで終えてしまったことは確かだ。
本来なら最終生物(LUCA)への移行過程の説明がくるはずだが、「原始ゲノム=自己」が「ゲノム<細胞=自己」という構造へと移行して、自己の範囲が拡大したとだけ述べてお茶を濁した。
しかしこれには理由がある。私はLUCAへの最終過程は、無生物から生物への過程より、生物のダーウィン進化のなかで理解したほうがいいと考えている。場合によってはダーウィン進化は今もゲノムの多様化と、選択の問題として語られる。しかし、ゲノムの発生以降、生物進化は様々な情報をゲノムの外に生み出してきたため、生物の進化をゲノムの進化としてだけ捉えることは難しくなった。私はこの問題は、原始ゲノムの代表する自己が、細胞を単位とする自己へと拡大した過程で最初に発生したと思っている。
原始ゲノムは現在の生物に普通に見られる様々なメカニズムにより情報を拡大させ、環境を自己に取り込むことでLUCAへの道筋を突き進んできた。しかし、生物の自己が完全に細胞に拡大してLUCAが誕生するためには、拡大したゲノム情報だけでは足りない。この困難は原核生物の細胞分裂過程をみるとよくわかる(図1)。
図1 原核細胞の細胞分裂調節と、LUCA以前のプロトセルとの比較
原核細胞のゲノム複製は、細胞が成長して一定の大きさに達したときに開始する。すなわち、原始ゲノムから見たとき拡大した自己=細胞の大きさを情報としてゲノムの複製開始が指令される。実際には細胞のサイズ情報を受けたdnaAと呼ばれる分子が複製開始点に集まり、ゲノムの複製が始まる。次にゲノムの複製が終わると、これが情報となって、細胞の中央がくびれ、細胞中隔が形成された後、細胞が分裂する。すなわち、今度はゲノムの状態が情報として細胞質の分裂を指令する。実際にこの分裂は、ゲノム複製に関する情報に反応してFtsZと呼ばれる分子が分裂面に集まることで進む。もし複製終了を情報として確認するメカニズムがなく、ゲノム複製が終わる前に細胞が分裂してしまったら大変だ。
この過程からわかるのは、原始ゲノムから細胞へと自己が拡大すると生物にとってゲノムの塩基配列だけが情報ではなくなり、複製開始を指令する細胞の大きさや、逆にゲノム複製の状態が情報となって細胞分裂過程の進行を調節していることだ。おそらく原始ゲノムは最初から脂肪膜でできたマイクロセルの中に存在してプロトセルを形成していたのではないかと考えられる。ただ、マイクロセル形成と、原始ゲノムは基本的には独立していたはずだ。(季刊生命誌89号に人工細胞研究の記事がある)細胞質・細胞膜とゲノムが独立しているプロトセルを考えるのは簡単だ。しかし自己が細胞として統合されるためには、原核細胞に見られるゲノムから細胞質、細胞質からゲノムへのシグナルが必要になる。すなわち細胞の大きさとゲノムの状態が、核酸配列情報の外に発生する必要がある。
「え?ゲノムの塩基配列以外の情報が発生したの?」と問われそうだ。答えは「イエス」だ。
ずいぶん前に、雨は情報でもなんでもない物理現象だが、外出しようと考えている人にとっては重要な情報として、傘を持って表に出るという行動につながることを例に、一見情報とは思えない現象が、解釈によって人間にとっては情報になることを説明した。まさにこの解釈システム(あるいは感覚システム)が原始ゲノムから細胞への拡大過程で発生することで、塩基配列以外の状態が情報として使うわれていることを意味している。
生命科学になじみのある人なら、今私が説明した機構はフィードバック、フィードフォワード機構として、生体の中では普通に見られるメカニズムだと気づかれるだろう。
生体ではこの機構が、細胞や個体の統合を保つための最も重要な機構として至るところで使われている。すなわち、生体内で起こるすべての独立の過程を統合するには、反応のモニタリングと、解釈、それに従う行動のネットワークが網目のように張られる必要がある。そして、それぞれの回路は、ゲノムからの指令で形成される分子により解釈される、ゲノムとは異なる情報が発生している。
もちろんSzotakらの研究が示すように、新しい情報解釈システムがなくとも、条件が整えばRNAゲノムを持つプロトセルが分裂を繰り返すことは現実にあり得る。しかし、この分裂でゲノムの分裂と細胞質・細胞膜の分裂がうまく調整されているように見えるとしても、それは地球上の偶然が重なりフィードバック機構を持った「間欠泉」ができるのと同じだ。図に示す様に、外界から何らかの力を受けることなく、ゲノムと協調して分裂することはない(Ottoの複製子を思い出して欲しい)。
この様に、原始ゲノムが誕生してから、生物は自己の範囲を急速に拡大し、これは自己内で起こる様々な現象を情報として解釈する無数の仕組みを獲得することで支えられてきた。例えば、タンパク質のリン酸化、アセチル化、メチル化、あるいはRNAのリボスウィッチなど、すべてこれにあたる。現在cAMPや脂肪酸がシグナル伝達に使われているのも、ATP、脂肪酸がこのとき細胞にふんだんに利用できた名残だろう。この再帰性が、目的性をもった力学系テレオダイナミック系を特徴付けていることを思い出して欲しい。
この様に新たな情報を解釈して自己の統合性を維持する機構が発展すると、今度は自己だけでなく外界の変化を情報として解釈するメカニズムも発生する。この結果、多細胞体制を支えるエピジェネティックな情報、神経回路情報、脳回路情報、そして最後に言語というそれぞれまったく異なる質の情報が生物から続々誕生してきた。
考えてみると、プロトセルの細胞質や細胞膜は、最初の原始ゲノムにとっては自己ではなく、環境だったと言える。この環境を取り込んで自己の範囲にしたのがLUCAの誕生と言えるが、これにより、その後の進化の方向性、すなわち環境の取り込み(支配)を目指す進化の原動力が決まった気がする。そして何よりも、その過程で例えばゲノム情報の多様化だけではなく、様々な異なる質の情報をゲノムの外に誕生させた。言語を考えてみると、このおかげで自己の範囲は世界の裏側、宇宙の果て、そして生命誕生のときまで広がっている。
これから生物の進化を、これら多様な情報が統合された個体を選択する過程として捉える必要がある。生物の進化をゲノム進化からだけ考えることはできない。次回からは、多様な情報が統合された生物の進化について考えてみたいが、まず生命の進化によって新たにゲノムの外に生まれた情報をリストし、その特徴を考えるところから始めたい。
[ 西川 伸一 ]
ゲノムを考える
2015年5月15日
序:ゲノムは情報
今はその気になれば全ての生物のゲノム解読が可能で、順を追って全ての種のゲノムを明らかにする国際プロジェクトが進んでいる。また個人レベルの進化研究も、ゲノムプロジェクトのおかげで大きく進展した。事実これまで30話にわたって紹介した論文のほとんどは、ゲノム解読を最も重要な研究手段として使っている。例えば昨年8月に第13話で紹介した何十羽のカラスのゲノムを比べる研究が行われる日が来るなど、私が大学を卒業した時には想像できなかった。誰もが同じ理解を共有していると思い込み、日常深い考えもなく頻繁に使っている「ゲノム」という言葉も、改めてその意味はなにか?と問われると答えるのは簡単ではない。そこで、30話を終えたところで少し立ち止まって、ゲノムとは何かについて何回かに分けて整理することにした。
いうまでもなくゲノムとは個々の細胞の中にあるDNA全体が代表している何かだ。DNA自体は物質の総称で、4種類の異なる塩基を持つヌクレチドがつながった巨大分子と定義することができる。しかし、ゲノムが持つ意味はこの物質性とは全く別のところにある。例えばゲノムは、我々が遺伝子と呼ぶものに対応する部分を持つが、遺伝子という言葉は、それがコードしている特定のペプチドや機能的RNAと対応して初めて意味を持つ。ヘモグロビンの遺伝子、ケラチンの遺伝子という具合だ。しかし、遺伝子=DNAではない。普通ヘモグロビンのDNAとは言わない(そう呼ぶ場合はDNAを遺伝子と同義に使っている)。これはDNAが物質で、遺伝子が機能をさす言葉だからだ。逆に言うと、遺伝子という概念には核酸という物質性は含まれる必要はないが(PC上に遺伝子を保存できる)、それが参照する分子の概念は必ず含まれる。ここでとりあえず遺伝子という言葉を、「DNAを媒体とする、特定の分子を参照する情報」と定義しておこう。
では情報としてのゲノムの性質を次に考えてみよう。遺伝子が情報として働くことを可能にしているのが、DNAの持つ情報媒体としての性質、即ち異なる塩基を持つ4種類のヌクレオチドが様々な配列で並ぶポリマーを作り、この並び方のパターンで無限のモノやコトを参照できるという性質だ。この情報は一定の法則にしたがって、実際の機能分子へと変換され生命を支える。また、AはT、CはGと相補的に対応することで自らの複製、伝達、転写が可能になる。見れば見るほど完璧な情報媒体だ。しかし、このDNAが持つ情報媒体としての完璧性が、ゲノムの理解を難しくしている。ゲノムの概念に何が含まれているか見てみよう。まずタンパクや機能的RNAへと転換される遺伝子が存在する。加えて、遺伝子発現制御やスプライシング制御など ‘コト’についての情報も同じようにA,T,C,Gの配列で表現されている。ただ問題は、核酸の並びにモノや機能を対応させられない配列、その機能や意味が全くわからない配列が、対応が付いている配列の何倍も存在している。この理解できない配列を情報と考えていいのか?存在しないのと同じと無視していいのか?これが決められない以上はゲノムも理解できない。繰り返すが情報媒体としてDNAは完璧だ。そのためDNAには必ず情報が存在すると思ってしまう。このことは文字と比べてみるとよくわかる。最初から文字は言語という情報の媒体として生まれてきた。従って、文字があるとそこに必ず情報があると私たちは確信する。だからこそ、初めて見たシュメール人の楔形文字も解読できると確信した。しかし例えばアルファベットをランダムに並べることは簡単で、いくら文字の並びが目の前にあったとしても情報を担っているかどうかはわからない。DNAも文字と同じで、その機能は情報をコードする媒体だ。このため、DNAには情報が必ず担われていると思ってしまう。しかし意味のない単語からわかるように情報媒体を使っても情報でない例はいくらもある。では文字と同じように、意味が理解できない配列は情報ではいと無視すればいいだろうか?実はそう簡単ではない。今回は短く触れるだけにするが、ゲノムには言語と異なるさらに厄介な問題がある。即ち情報の出し手がいないという問題だ。言語の場合、情報には必ず出し手と受け手が存在する。一方、ゲノム上の情報には出し手はいない。シャノンの情報理論を見ると、出し手がいるということで100%正確な情報がまず存在することが前提になっている。即ち何が情報で、何が情報でないかは最初から決まっている。一方、ゲノム上の核酸配列は、何が情報で、何が情報でないか最初からは決まっていなかった。というより、そもそもゲノムには情報としての出し手がいない。誤解を恐れずいうとDNAという情報媒体は自ら新しい情報を生み出せるという性質を持つ。このため、今私たちが情報でないと判断した部分も、将来情報化できるポテンシャルを持っている。さらに一つのゲノムが新しい情報を生みだすのに一部の部分だけが関わるわけではなく、ゲノム全体が様々な形で新しい情報の誕生に影響を及ぼしている可能性がある。即ち、部分と全体が常に一体化していて、分けることができないという生物の特徴的関係が、ゲノムレベルにも存在するようだ。従って、ゲノムは部分として様々な情報をコードするだけでなく、全体として新しい情報の誕生や消滅に関わっている可能性が高い。このようなゲノムの性質は、出し手のはっきりしたシャノン情報理論でゲノムを捉えることが難しいことを意味している。このように、情報としてゲノムが持つ数々の問題がある。これから「ゲノムを考える」で、この問題をもう少し深く掘り下げながら、ゲノム理解を妨げている問題を整理したいと思っている。
「ゲノムを考える」導入部、第一回の結論は「ゲノムは情報」だ。このことを示した面白い論文を一つ紹介して31話を終わる。山梨大学教授の若山さんがCDB(理化学研究所 発生・再生科学総合研究センター)在籍時に行った仕事だ。昨年は小保方問題の渦中の人になってしまった若山さんのライフワークは絶滅種、あるいは絶滅危惧種をクローン技術で復活させることだ。この模擬実験として、若山さんは16年フリーザーの中に捨てられていたマウスの死体の細胞から核を取り出し、核を抜き取った未受精卵に移植した。無論16年経っていると言ってもひょっとして生きた細胞が残っているかもしれない。核移植に使った細胞が全部死んでいることを確かめるため、フローサイトメーターで100万以上の細胞を調べても生きた細胞を見つけることができなかった。このように完全に死んでいることを確認した細胞から核を取り出し、卵に移植すると高い確率で胚発生が始まり、その胚から多能性幹細胞株(ES細胞)を樹立することができた。さらに、このES細胞株をマウス胚盤胞に移植して発生させると生殖細胞に分化し、ついに死んだ細胞の核から生きたマウスを作成することに成功した。若山さん自身は論文の中でこの結果をもっぱら技術的進歩として議論しているが、私にとってはゲノムが情報であることの最も明確な証明に見える。すなわちゲノムの完全性は細胞の生き死にに関わらず維持されている。情報の完全性に、生命というシステムは必要ない。情報を実験的に扱うときは、物質として扱えばいい。この実験系なら、削ったり、足したり、再編成したゲノムからクローンを作ることで、ゲノム情報の全体と部分の関係を問い直す実験ができるかもしれない。
[ 西川 伸一 ]
ゲノムの解剖学
2015年6月2日
31話でゲノムは情報だと結論した。では私たちのゲノムがどんな要素からできているのか、まず解剖学から始めよう。実際にはミトコンドリアにもゲノムはあるのだが、話を簡単にするために核内のゲノムに限る。一部の細胞(例えば赤血球)を除いて、私たちの細胞の全てに核があるが、ゲノムはこの核内にあるDNA総体が表現している情報だ。核内には46本に分かれたDNAが存在し、ヒストンをはじめとする多くのタンパク質と結合して、コンパクトに折りたたまれ、染色体を形成している(図1)。この染色体一本一本の基本的構造は同じなので、ここからはゲノムが一本の染色体からできているかのように話を進める。
図1 ヒトのゲノムは核の中に46本の染色体に分かれたDNA。
ヒトのゲノム全体は、約30億個のヌクレオチドが連なった高分子で、ヌクレオチドの持つ4種類の塩基の組み合わせが媒体となって全ての情報が書かれている。ゲノム内の情報がコードする最も重要なものが、細胞という化学反応系を形成・維持するための基本分子であるたんぱく質で、これに対応するゲノム部分をcoding DNAと呼んでいる。何をcodingと呼ぶかは定義の問題なので、ここではタンパク質まで翻訳される遺伝子をcodingDNAと呼ぶ。ただこのcoding DNAに対応する遺伝子はヒトで22000程度しかなく、全ゲノムに占める割合は1.5%もない。少数の例外を除いて、個々のたんぱく質をコードする遺伝子はエクソンと呼ばれる小さな部分に別れて存在している。一度RNAに転写されてから、たんぱく質の情報に関わらない部分を除去した一本のmRNAへとつなぎ合わせた後翻訳される。このエクソンとエクソンの間にある配列をイントロンと呼ぶ。遺伝子が情報としての機能を発揮するためには、他にも遺伝子を正しい場所からRNAに転写するための様々な指令情報が必要だ。このほとんどは、エクソンの外、すなわちイントロンや遺伝子から様々な距離で離れたnon-coding領域に存在している。これからわかるのは(図2)、たんぱく質という「モノ」に対する情報(エクソン)と、モノの情報を書き出すための様々な標識、すなわち「コト」に対する情報から一つのcoding単位ができていることだ。
図2 coding遺伝子単位の概略。調節領域の指令で遺伝子が転写されると、RNA maturation に関わる標識により、イントロンが除去された一本のmRNAが形成される。
「モノ」「コト」両方の情報を含めると、coding単位がゲノムに占める割合はずっと大きくなるが、逆にその正確なサイズは測定しにくくなる。なぜなら、「コト」に必要な情報が、ゲノムのどこからどこまで必要かを正確に算定できないからだ(これは次回に議論する)。さらに、同じ遺伝子領域に逆向きに異なる遺伝子が並んでいることもある。したがって、正確にcoding遺伝子セットが占めるゲノム上の割合を算定することは難しい。
次にcodingとは言えないが、構造的には遺伝子と呼べる単位が存在している。ただ、正常な遺伝子とは異なり、そのまま翻訳されタンパク質ができることはない。そのため、偽遺伝子と呼ばれている。ヒトでは偽遺伝子と考えられる部分がなんと1万以上も存在している。進化の過程では、coding遺伝子が重複を繰り返して多様性を拡大するのをよく見ることができる。このように重複した遺伝子に変異が導入されて機能がなくなると偽遺伝子と呼ばれるようになる。偽遺伝子が最も多いのは嗅覚受容体で、ヒトでは400の機能的遺伝子に対してなんと600もの偽遺伝子が存在している。ただ、偽遺伝子だからなんの役割もないと決め付けることはできない。成り立ちから考えても、偽遺伝子といえども、構造的には遺伝子と同じだ。しかも、機能していないことから、変異が蓄積されやすく、新しい機能を持った遺伝子を作るという目的にはうってつけの材料だ(例えば第21話参照)。この意味で、遠い将来、新しい機能を持つ分子の資源としての役割を担っているということができる(図3)。また、ニワトリの免疫グロブリン遺伝子で見られるように、機能的抗体遺伝子と遺伝子転換により部分の交換を行うことで抗体の多様性を生みだすための鋳型として働いている場合もある。このように、ゲノム各領域を、それが必要とされる時間スケールを変えて見ることは、ゲノム理解について重要で、これについても次回以降議論する予定だ。
DNAにコード可能な生体機能分子はタンパクだけではない。これまで多くの機能を持つRNA分子が知られている。リボゾームRNAやトランスファーRNAなどのタンパク翻訳に関わるRNAはその代表的なものだが、他にもスプライシングやRNA編集などのRNA成熟、DNA合成、遺伝子制御、トランスポゾン制御など、多岐にわたる機能を持つRNAが知られている。機能の詳細は省いて図にまとめておくが、機能的RNAはゲノムの4%ぐらいを占めていると考えられている。これらを一つ一つ紹介するつもりはないが、この中のnoncodingRNAは、DNAが持つもう一つの顔を示していると考えており、これも次回以降議論する。

図4 様々な機能的RNA このリストは機能的RNAの一部。
ここまで主に転写される部分について見てきたが、ゲノムには多くの転写されない場所が存在する。例えば、染色体のヘテロクロマチンと呼ばれる場所に存在する繰り返し配列の集合は、全ゲノムのおおよそ6%を占めている。分裂時に染色体を引っ張る微小管の結合部位である動原体が結合するために使われているセントロメアや、分裂時に染色体が削られて重要な遺伝子が消失するのを防いでいるテロメアなどは、転写はされなくてもゲノムの複製や維持に必須の領域であることがわかっている。しかし、ヘテロクロマチンを形成する他の多くの繰り返し配列は、機能がよくわかっていない。実際、ヘテロクロマチンとそれに対応する繰り返し配列は、種を越えてひろく存在しているが、その配列自身は保存されていない。他にも、超可変ミニサテライト(hypervariable minisatellites)と呼ばれる、ユークロマチンに存在する繰り返し配列も存在する。
ヒトゲノムの解読結果を見た時の一番大きな驚きは、トランスポゾンに由来する配列が全ゲノムの5割近くを占めていたという事実だ(図5)。トランスポゾンとは、基本的には細胞の中で染色体から染色体へ伝搬できる遺伝子単位を指し、図に示すようにLINE, SINE, LTR—トランスポゾン(レトロビールス型)、そしてDNAトランスポゾンに分類されている。
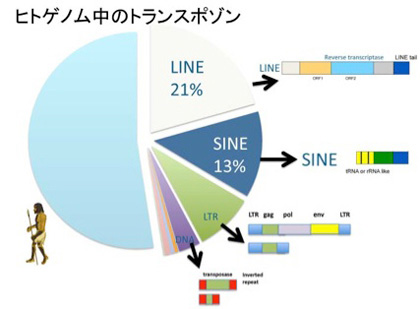
図5 ヒトゲノム中に見られるトランスポゾンの種類とゲノム上の割合。
これらはレトロトランスポゾンと、DNAトランスポゾンに大別され、レトロトランスポゾンはレトロウイルスと同じように、ゲノムからはRNAとして読み出され、そのRNAが逆転写酵素でDNAに転写されたあとゲノムに挿入されることで伝搬する。完全型のレトロトランスポゾンは、従って逆転写酵素をコードしている。逆転写酵素を持たない不完全型レトロトランスポゾンの場合は、完全型のトランスポゾンが発現する逆転写酵素を利用することで伝播が可能だが、基本的には伝搬能力を失っていると見たほうがいいだろう。RNAとしてゲノムからそのまま転写されるレトロトランスポゾンと異なり、DNA型トランスポゾンは自らを正確にゲノムから切り出し、また他の箇所に挿入するためのトランスポゼースと呼ばれる酵素をコードしている。多くの種では、完全なトランスポゼースを持ったトランスポゾンがゲノム中に存在しているが、ヒトゲノム中に存在しているほぼすべてのトランスポゾンは、トランスポゼースの活性を失っている。従ってトランスポゾンの活性を失った化石といえる。
今回は話を進めるための基礎知識として、ゲノムの構造要素の概略を述べた。こうしてヒトゲノムを解剖していくと、こじつけもあるが、おおよそ6割の配列になんらかの意味を対応させることができる。ただ意味や機能が想像だにできない部分が4割近く存在する。また私たちのゲノムのほぼ9割をトランスポゾンか、意味のわからない配列が占め、これらの塩基配列に何か積極的な意味があるようには思えない。このため、ジャンクDNAと呼ばれたりしている。ただ、意味が無いのか、理解できないだけなのか、9割を占めるジャンクについて考えることが必要だ。もちろん種によって、この構造は多様で、ジャンク配列のゲノムに対する割合も大きく違う。次回からは、ゲノムを構造化することで新しい情報が生まれるようすを見ながら、さらにゲノムの理解を進めてみたい。
[ 西川 伸一 ]
構造としてのゲノム
2015年6月15日
ここまで、ゲノムが細胞の生き死にに左右されない(言って見ればPCが止まっていてもメモリーは残るようなもの)情報であることを見てきた。しかし、ゲノムに組み込まれた情報が全てATCG塩基配列で表現されていると考えると、大きな間違いを犯すように思う。すでに見たように、翻訳されるたんぱく質をコードする情報自身は塩基配列で表現されているが、ゲノム上に全くランダムに存在するのではなく、適当な長さのエクソンに分かれ、それを転写するための様々な指令情報が組み合わさった単位として存在している。すなわち、一定の構造をとっている。「何を今更?」と言わずに続いて聞いてほしい。この転写を調節する時に使われる様々な「コト」についての指令標識は塩基配列として表現されるが、それ以外にも情報化されていることがある。例えばイントロンの長さ、エンハンサーの位置などは、ゲノム上の構造として表現されるものだ。もしイントロンの長さが遺伝子の転写に重要な役割があるとすると、全く意味がわからない塩基配列も情報として意味を持つ可能性がある。我々のゲノムのかなりの部分が、情報として具体的な何かに全く対応していないジャンクと呼ばれるDNA配列だと述べたが、これも塩基配列で表現できる情報としてみると意味がわからないからジャンクと呼んでいるだけで、構造としてゲノムを見た時重要な役割を持つのかもしれない?そこで、まずゲノムが構造化されることで生まれる塩基配列とは違った情報の可能性について見ていこう。
1、遺伝子の秩序立った配置:Hox遺伝子の衝撃
ゲノムが構造化されることで、DNAが新しい情報媒体になることを我々が最も実感したのは、頭から尻尾まで、体の各体節のプランを決めているHox遺伝子のゲノム上の構造が明らかにされた時ではないだろうか(この遺伝子の変異で触覚が足に変わったショウジョウバエについては既に第6話で述べた)。急速に進む様々な種のゲノム解析から、ほぼ全ての動物種で、何個ものHox遺伝子が小さな領域にクラスターを作って集まっていることがわかってきた。図1に示すように、現在ではこのクラスター内のHox遺伝子を5’から順にHox1,Hox2….Hox13と名付けている。
図1ショウジョウバエと哺乳動物のHox遺伝子構成と、その発現。体軸に沿った発現のパターンが、遺伝子の構成と対応している。この対応関係はcollinealityと呼ばれている。
脊椎動物より未熟な動物ではこのクラスターは一つしかないが、脊椎動物では重複により別々の染色体に4つのHox遺伝子クラスターが存在する。驚くべきことに、体の体制が完成した胎児期にHox遺伝子の発現を調べると、ショウジョウバエも、マウスでも図1に示すようにゲノムに並んだ順番、すなわちHox1からHox13まで順番にそって、頭部から尾部へHox遺伝子が発現している。このゲノム上の遺伝子の配置と、頭部から尾部への発現場所の対応性はcollinealityと呼ばれ、左右相称動物では広く保存されており、このHox遺伝子の決まった配置を維持することの重要性を示している。
図2 Hox遺伝子クラスターの進化。四足類ではHoxAからHoxDまでの4クラスターが存在するが、便宜的にこの図ではHoxAクラスターで代表させている。
次に、図2に従ってHox遺伝子クラスターの進化過程を見てみよう。第一段階では遺伝子重複によりまず4種類のHox遺伝子が並んだクラスターが形成される。こうして生まれたゲノム上のHox遺伝子クラスターは、次の左右相称動物の進化過程で原型が完成し、各Hox遺伝子の塩基配列だけでなく、配置そのものが情報として体の前後の体制の形態形成に利用されるようになる。その後それぞれの動物の体制に合わせてHox遺伝子の数は増減し、また配置も変化しているが、基本的には各遺伝子の配置は前後軸に合わせて構造化されたまま維持される。この事実を見ると、遺伝子の配置という構造が塩基配列とは異なる情報をゲノムの中に持ち込んでいるのがわかる。構造やデザインが機能を持ち、情報になることは生物の世界では当たり前だ。当然ゲノムも形質と同じように、特定のデザインの下に構造化されることで新しい情報を持っても不思議はない。例えばHox遺伝子の塩基配列はそのままで、順序が狂うことで新しい体の体制が生まれたとしよう(例えば尾が長くなる)。もしこの結果生まれた形質が自然選択されたとすると、この時選ばれた変異は塩基配列の変異ではなく、遺伝子の配置、すなわち構造の変異が情報として選択されたことになる。あるいはまたHox遺伝子間の距離が一定に保たれることが重要だとすると、その距離を埋めているのは特定の配列というより、ATCGを積み重ねて一定の長さを実現できるDNA の物理的性質によっていると言える。すなわち、塩基配列が問題ではなく、構造が問題になっている。重要なことは、構造化により生まれる情報も、子孫に伝える時は塩基配列情報として複製される点だ。複製だけを見ていると塩基配列だけが情報だと勘違いするが、このように伝達時には塩基配列情報として、機能発揮時には構造情報として働くゲノム部分があることを無視してゲノムは考えられない。
2、ヌクレオソーム
染色体を見ると、塩基配列とは無関係にDNAを基材とする構造が形成されている。これもDNAが持つ構造化能力の例だ。具体的に見てみよう。図3に示すようにDNAは4種類のヒストンが集まった8量体に巻きつくヌクレオソームという構造を取っているが、ヒストンとDNAの結合には原則として特異的な遺伝子配列は必要ない。
図3 ヌクレオソーム単位の構造。一つのヌクレオソームには200bpのDNAが巻きついている。
<引用>
上図:http://commons.wikimedia.org/wiki/File:Nucleosome_structure_and_subunits.svg
下図:http://commons.wikimedia.org/wiki/File:Pioneer_Factor_rearrange_the_nucleosome.jpg
これまでヌクレオソーム構造は、長いDNAを核内にコンパクトにたたむために進化してきたと考えられてきた。しかし、遺伝子の大きさが4Mbしかない古細菌類の一部がヌクレオソームをもつことが発見され、今では転写を効率化するために進化してきたと考えるようになっている。実際、古細菌、真核生物の両方で、転写開始点のほぼ全てがヒストンに巻きついていないリンカーDNA部位に存在している。すなわち、ヌクレオソームは高々200bpの長さのDNAからなる小さな単位で、構造も単純なため複雑な情報を持つことはないが、ゲノム内の最小構造単位として、転写開始場所を構造的に指示するために進化してきたようだ。
ヌクレオソームの出現は、その後の生物情報の可能性を一変させ、最終的に人間の言語獲得にまで至る道を決定付けたと思っているが、このことはまたいつか議論しよう。ともかくヌクレオソーム構造獲得のおかげで、このコアヒストン構造をさらに修飾してゲノムの使い方を決めるエピジェネティック機構が進化し、ゲノムにさらに複雑な構造化を導入することを可能にした。実際先にあげたHox遺伝子の配置はこの新しい機構を巧みに取り込んでいる。体の節、すなわち体節を形成する中胚葉では最初HoxA遺伝子クラスターは全てH3ヒストンの27番目のリジンがメチル基で修飾されており、これによりHoxクラスター全体の遺伝子の発現が抑えられている。即ち、ヒストンの修飾により転写に必要な分子が近づけない領域を構造的に保証している。この抑制型ヒストンは発生が進み前方から体節が発生する時には、前側のHox遺伝子領域から外れ、体の体制に合わせてHoxクラスターが発現できるための重要なメカニズムになっている。最終的に、体幹部では後ろ側のHox遺伝子が存在する領域は抑制型ヒストンが維持されるため、後ろ側のHox遺伝子は発現できない。一方、体の後方ではほとんどの領域から抑制型ヒストンが外される。このようにHoxクラスターの配置が体の位置と対応しているおかげで、Hoxクラスターの発現調節を大まかに区域化することが可能になり、体の複雑なプランを指示することが可能になっている。ただ、H3K27のメチル化を介するゲノムの区域化は特殊な例で、普通はH3の4番目のリジンがメチル化したH3K4me3と、9番目のリジンがメチル化したH3K9me3が引き金となって、染色体構造をそれぞれユークロマチンと呼ばれる開いた状態と、ヘテロクロマチンと呼ばれる閉じた状態に構造化することが、最も普通に見られる転写活性の区域化のやり方だ。この構造化はDNA自体のメチル化も含め様々な分子が関わる複雑な過程で、マトリックスとしての ゲノムを考える時に議論したい。また、生命誌ジャーナルでもクロマチンについて国立遺伝研究所の広瀬さん、前島さんに解説をいただいているので、是非参照いただきたい
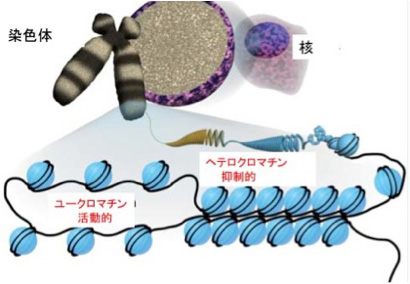
図4 クロマチン構造。染色体を見ると、ヌクレオソームの密度が高い場所(ヘテロクロマチン)と、低い場所(ユークロマチン)を区別することができる。この密度も、ヒストンの異なるメチル化を引き金にゲノムが構造化された現れで、ヘテロクロマチン領域での転写は抑制されている。
引用<Sha, K. and Boyer, L. A. The chromatin signature of pluripotent cells (May 31, 2009), StemBook, ed. The Stem Cell Research Community, StemBook, doi/10.3824/stembook.1.45.1. http://www.stembook.org/node/585>
今回は、ヌクレオソームを基盤として、ゲノムを構造化する重要な機構がエピジェネティックス機構だという結論で止めておく。ただ、ヌクレオソームやエピジェネティック機構の進化は実際にはHoxクラスターの進化より先に起こっている。したがって、エピジェネティック機構というゲノム構造化を可能にする機構をさらにうまく利用する方法の一つとして、Hoxクラスターのような新たな構造化が加わってきたと考えるのが正しい。次回は、ヌクレオソームより大きなゲノムの構造単位、topology associating domaiから話を続ける。
[ 西川 伸一 ]
ゲノムの構造単位
2015年7月1日
1、topology associating domain (TAD)
最近、遺伝子の組織や発生時期特異的な発現調節の基本構造単位としてtopology associating domain(TAD)と呼ばれる領域に注目が集まっている。ヒトゲノムには約2000のTADが存在すると考えられているが、TADの存在が広く認められるようになったのはつい最近のことだ。TAD概念の確立には、核内で位相的に接して存在しているゲノム領域を特定するchromosome conformation captureと呼ばれる方法の開発が大きく寄与している。一つのTADは平均500kb—1Mb程度の大きさで、その中に1個から複数の細胞特異的、あるいは発生時期特異的に働くcoding遺伝子と、その発現調節(エンハンサー)領域を含む長いDNA単位が大きな塊を作って核内に存在していると考えられる。図1はPopeらの論文(Nature, 515:402, 2014)から拝借したTADのイメージ図だが、それぞれのTAD単位を構成するDNAが、分離した毛糸玉のようにまとまって核内に存在していることが表現されている。なぜこのような構造化が必要なのか?これについては、遺伝子発現の時期や場所を決めている調節ユニットを一つの塊として構造化する必要に答えたのがTADではないかと考えられている。
図1 TADの概念図と、HiCヒートマップ:説明は本文参照
実際、HiCと呼ばれるChromosome conformation capture法を用いて、どの領域がどの領域と隣接していかを調べると、遺伝子やエンハンサーの相互作用がTAD内に制限されていることがわかる。図1のヒートマップはHiCの結果をマッピングしたもので、各領域がコンタクトしている確率が高いほど赤くなるよう示されている。核という狭い3次元空間では、原理的に各領域はどの領域とも近接することは可能だが、実際にはTAD内にある領域は同じTAD内の領域と相互作用する確率が高い。TAD内部の塩基配列は特に保存されているわけではないが、各TADの間には、種間でよく保存された、CTCF分子の結合配列、house keeping遺伝子、tRNA遺伝子、そしてSINEトランスポゾンなどが集まった境界領域が存在している。最近の研究から、この境界が、一つのTAD内のエンハンサーの効果が、隣接したTADに影響を及ぼさないよう制限するインシュレーターの働きをしているのではと考えられるようになってきた。私たちの体は何百種類もの細胞から出来ており、それぞれの細胞ごとに働いている遺伝子は違っている。とはいえ、それぞれの細胞分化に必要な遺伝子群を、分化ステージや細胞系列に合わせて狭い領域にまとめることは難しい。このため全く異なる細胞で発現する遺伝子が隣接して存在することはゲノムでは普通に見られる。従って、一つのTADの発現が、隣のTADにある遺伝子の発現に影響を及ぼさないよう構造化されていることは重要だ。次に、これをより具体的に示した2編の論文を紹介しよう。
論文1:TADによるHox遺伝子制御(Andrey et al, Science 340, 1234167, 2013)
TADによるゲノム構造化の重要性が強く認識されたのもまたHox遺伝子の研究からだった。HoxDクラスターは腕と指両方の発生に関わっているが、例えばHoxD10のマウス発生時の発現を経時的に調べると、図2に示すように、四肢発生の初期段階で前腕の形態形成に関わっていたHoxDクラスターが(図2:青色で示した部位)、時間とともに今度は指の形態形成に関わるようになるのがわかる(図2:茶色で示した部位)。同じHoxクラスターがほとんど同じ発生時期と場所で異なる構造の発生に順番に関わることを可能にするメカニズムを追求していたスイスのDubouleらは、HoxDクラスターが2つの隣接するTADの境界に存在することを見出した(図2)。
図2 Andrey et al, Science 340, 1234167,
2013 論文解説図。
HoxDクラスター内の遺伝子が、前腕から手指へと発現が移動するのに呼応して発現を調節するエンハンサーも、TAD2のエンハンサーから、TAD1内のエンハンサーに移行する。
さらに、それぞれのTADに存在するエンハンサーの活性を調べると、テロメア側のTADには前腕での発現に関わるエンハンサーが、セントロメア側のTADには手指で発現するエンハンサーが集中していることをつきとめた。この結果からDubouleたちは、HoxDクラスターの全ては最初テロメア側のTADの支配を受け前腕形成に関わっているが、発生が進むと一部がセントロメア側のTADの影響を受けるようになり、徐々に支配されるTADをシフトさせることで、前腕と手指の発生に連続的に関わることを示した。すなわち、もともと構造化されているHoxクラスターに、TADという遺伝子調節単位構造が絡むことで、発生時のHoxD遺伝子発現の複雑なパターンが実現していることを示した。
論文2:TAD境界領域の機能(Lupianez et al, Cell 161, 1012, 2015)
Dubouleの研究からTADが構造化されたエンハンサーの供給単位になっていることがわかるが、TAD同士が影響し合わないよう機能的に分離されているかどうかはこの実験からはわからない。最近ドイツベルリンから、遺伝子領域の大きな欠失、反転、重複によりTADの境界が崩壊すると、TAD内のエンハンサー領域の影響が隣接するTADに拡大して、その結果発生異常が起こることを示す重要な論文が発表された。TAD間の境界の機能を教えてくれる研究なので、図3を用いて紹介しよう。
図3 Lupianez et al, Cell 161, 1012, 2015 論文の解説図:説明は本文参照
この研究では手指の発生異常に関わる変異が集中するヒト第2染色体の約2Mbの大きさの領域に注目して、まずそれぞれの奇形がどのような変異から起こっているのか調べている。図3に示すように、この領域には3つのTADが存在し、それぞれのTADには様々な組織の発生に関わる増殖因子や転写因子をコードする遺伝子が存在していることが分かった。これら遺伝子のうち、手指発生で発現が見られるのはTAD2に存在するEPHA4(ephrinに対する受容体)だけで、TAD1,TAD3にある遺伝子は手指には発現しない。このことから、手指発生時に遺伝子発現を誘導するエンハンサーの効果がTAD2内の遺伝子のみに限局されていることがわかる。次に、手指発生異常をおこす遺伝子変異を解析すると、境界2として示した領域が大きく欠損する変異では指が短くなる短指症が、境界1を含む領域の逆位が起こると、親指と人差し指の融合(F-syndrome)が、同じく境界1を含む部分の欠損では多指症が発症することが分かった。同じ変異を今度はマウスゲノムに導入して、人間とほぼ同一の手指発症異常をマウスで再現できることを確かめ、それ以降の実験はマウスで行っている。マウス手指発生過程でTAD1,TAD3に存在する3種類の遺伝子の発現を調べると、境界2がなくなるとTAD2エンハンサー活性がPax3遺伝子に及び、手指発生でこの遺伝子が間違って発現してしまう。同じように、境界1が変異を起こすと、手指発生時にIHHやWnt6の間違った発現が見られることが分かった(図3)。この結果は、核という極めて狭い領域に閉じ込められ3次元的に絡み合ったDNA上の遺伝子が一定の時と場所で正確に発現するために、TAD間の境界が重要な機能を持っていることを示している。しかし、この境界がなぜTAD外のエンハンサーの影響を食い止められるかについてはよくわかっていない。おそらく、さらに大きなレベルの、ゲノム構造化と関わるのではと考えられるが、これについては次回に紹介しよう。TADを説明するため、今回は手指発生に関わる遺伝子の発現についての研究した2つの論文を紹介した。最後に少し余談になるが、手指の発生異常についての詳しい研究を最初に行ったフランス人Maupertuis(図4)を紹介して今稿を終わりたい。

図4 Maupertuis肖像(wikipediaより)
Maupertuisは、地球が縦にひしゃげた偏円であることを証明するためフランス科学アカデミーがペルーとラップランドの2カ所で観測を行った際のラップランド隊を率いた業績で有名になった物理学者・数学者だが、ビュフォンとともにフランス自然史を支える中心人物でもあった。彼はデカルトの機械論とそれを支持する前生説に反対し、後生説的有機体論展開の必要性を唱えた。1744年プロシア皇帝フリードリッヒ2世に強く請われてMaupertuisはプロシア王立科学アカデミーの会長に就任する。例えてみれば、日本学術会議の議長に中国や韓国から人を招くのと同じことで、待ち構えていたベルリン科学会の大きな抵抗にあうが、王室の後ろ盾と数学者オイラーの助けを借りて様々な改革を断行した。動物発生についてのMaupertuisの考えは、ライプニッツのモナド(単子)の概念に近い粒子が後生的に構造化されるというものだった。当時最も有力な説だったデカルトの機械論に基づく前生説を論破する根拠を探していた時、ベルリンで4世代にわたって多指症が維持されているRuhe家を知る。同じ形態異常が一族の母親からも、父親からも子孫に伝えることができることは前生説を否定する現象だとして詳しい研究を残している。ゲノム構造化単位TAD の意義を明らかにする記念すべき論文が、手指発生異常研究で最も長い歴史を持つベルリンからの論文であることを知り、Maupertuisを思い出し、深い感慨にかられた。
[ 西川 伸一 ]
核によるゲノムの構造化
2015年7月15日
A対T、 C対Gの結合をベースにした2重らせん状の分子構造と、4塩基の並びで表現できる情報の無限性を考えると、DNAほど情報媒体として優れた物質はちょっと思いつかない。この線状に並んだ塩基対が持つ情報としての可能性に圧倒され、塩基配列を線状のコードとしてだけ見てしまうと、ゲノムが2次元、3次元と構造化されていることを忘れがちになる。しかし、遺伝子の配置、ヌクレオソーム形成、クロマチン構造、そしてtopology associating domains(TAD)と見てくると、ゲノムが1次元の配列情報だけでなく、構造化することで新たな情報を担えることを理解してもらえたのではないだろうか。ただ、構造化はこれだけにとどまらない。ミトコンドリアDNAを除くと我々のゲノムの全ては核の中に折りたたまれて存在している。今回はゲノムが収納されている核の構造とゲノムの関係について見てみたい。
クリエーティブコモンズに登録されている画像をchromosomal paintingで検索すると、Robert MayerらがBMC Cell Biology 6:44, 2005に発表した論文を見つけることができる。以前私の研究室に在籍していたTimm Schroeder君も著者に加わっているのを発見し、意外な出会いに喜んだが、この論文では染色体の核内の位置がランダムに決まっているわけではなく、細胞の種類に応じて特定の位置に収まっていることを示した論文だ(図1)。この写真に見るような染色体の核内での位置はクロモゾームテリトリーと呼ばれている。
図1ES細胞(上図)とマクロファージ(下図)の1番と14番(左図)、あるいは2番、9番(右図)染色体をin situ hybridizationにより染めだしている。(Mayer et al, BMC Cell Biology 6:44, 2005)
図1は各染色体が、細胞の種類に応じて異なるクロモゾームテリトリーを形成していることを教えてくれる。すなわち、全体(核)が部分(染色体)の単純な集まりではなく、部分(染色体)が全体(核)によっても組織化される生命特有の特徴が、核と染色体の関係にも言えることを示している。もちろんクロモゾームテリトリーが染色体の位置を完全に決めているわけではない。細胞が分化して特定の遺伝子の発現が必要になると、その遺伝子が位置する領域はクロモゾームテリトリーから飛び出してくることも知られている。例えば、これまで幾度も登場したHoxD遺伝子の核内の位置をFISHと呼ばれる方法で調べた研究では、ES細胞分化に伴いクロモゾームテリトリー内に収まっていたHoxDがクロモゾームテリトリーから外れて存在することがわかっている(Morey et al, Development 134:909, 2007)。核全体から見たときの、このようなゲノム構造化を調節する仕組みはよくわかっていないが、他のゲノム領域と同様、次世代シークエンサーの開発により、核内に存在する構造とゲノムとの関係を直接調べることが可能になった結果、この分野は大きく進展しつつある。単純に見える核も複雑な構造を持ち、核酸以外にも多くの分子がこの構造を支えている。したがって、核によるゲノムの構造化についての研究は、ゲノムがコンタクトしている分子や構造の数だけ存在することになる。これではきりがないので、ここでは研究が進んでいるlamina associated domain(LAD)とnucleolus associated domain (NAD)について簡単に見ておこう。
・LAD(lamina associated domains)
図2 核膜直下の構築。青く染まるのは核酸、赤く染まるのは核膜孔。ここではLaminBを示しているが、核膜直下はLaminA,
LaminBが寄り合わさったラミナにより裏打ちされている。
(Wiki Commonsより)
電子顕微鏡を用いた核の解析から、核膜直下にはヘテロクロマチン構造が多いことは古くから知られており、このことから核膜直下に位置する遺伝子の発現は抑制されているのではないかと想像されていた。核膜はLaminA,Bを主成分とするラミナに裏打ちされている(図2)。ラミナとコンタクトしているDNAを標識するDNA adenine methyltransferase identifyication(DamID)と呼ばれる方法が開発され、これによってマップされる領域をlamina associated domain (LAD)と呼んでいる。DamIDは哺乳動物には存在しないアデニンメチル化酵素を利用して特定のタンパクと接する DNAをメチル化標識する方法だ。LADを検出するときはラミナの主成分であるLaminB1にアデニンメチル化酵素を融合させた遺伝子を細胞に導入、この細胞のラミナとコンタクトを持つDNAのアデニンをメチル化させる。哺乳動物のDNAにはメチル化アデニンが全く存在していないため、この細胞のゲノム中のアデニンがメチル化された領域を特定することで、LADを定義することができる(図3)。
図3 LAD決定のためのDamID法。DamIDとLaminB1のキメラ遺伝子を作成して細胞に導入する(黄色の丸印)。DamIDの作用によりラミナとコンタクトしているゲノム領域のアデニンがメチル化される。これを抗体で精製して配列を決める。
これにより、ヒトやマウスでは0.1−10Mbの大きさのLADが1300程度存在し、全ゲノムの4割を占めることが分かった。LADはcoding遺伝子が少ない領域が濃縮されており、75%程度は細胞の種類を問わず保存されている。逆に言うと、残りの25%は細胞種に応じてLADだったり、なかったりしておりで、LADが発生によりダイナミックに変化する構造であることを意味している。LADに存在する領域に結合するヒストンの構造などと比較する研究から、LAD内の遺伝子は一般的に抑制されていると考えられている。最近の研究で、TADと領域が重なっている場合があること、あるいはCTCF (CCCTC-binding factor,抑制性の転写因子)がLADの境界に結合していることなどが明らかになり、両方の構造化はメカニズムを共有していることが想定されている。しかしLADの研究は始まったばかりで、TADやクロモゾームテリトリー、あるいは次に取り上げるNADとの関係や、ラミナとコンタクトをとるメカニズム、あるいは発生過程でダイナミックに組織化されるメカニズムなど、まだまだ研究が必要だ。とりあえずこれまで分かっていることを単純化して、図4にTAD,LADの関係が細胞分化で変化する様子を示しておく。
図4 細胞分化時にはLADの位置が変化することでTADの活動を変化させる。一般的にTADの境界はLADとオーバーラップすることが多い。LADの影響を受けるTADはより強くラミナの影響を受ける。
・Nucleolus associated domain (NAD)
核小体はリボゾームRNAが転写され、リボゾームが形成される核内の一種の構造物だ。この構造とコンタクトしているゲノム領域を網羅的に調べる研究も行われ、NADと呼ばれている(図5)。
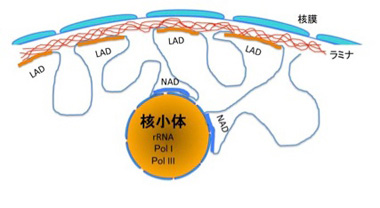
図5 LADとNADの位置関係。NADは核小体とともに精製されるゲノム領域を指す。
NAD領域の特定は、核小体と共に精製されてくるDNAを採取し、次世代シークエンサーで調べることで行われる。従来、核小体にはRNAポリメラーゼのうち、Pol I, Pol IIIが選択的に存在していることが分かっていたが、NAD領域にも、Pol I, Pol IIIによって転写される遺伝子が濃縮していた。このことから、確かに核小体の構造と密接に関わっているDNAが濃縮されていることがわかる。ゲノム全体の40%近くを占めるLADと比べると、NAD領域ははるかに小さく、ゲノム全体の5%に満たない。もっともはっきりしたNADの特徴は、染色体分配や細胞分裂時に重要な細胞小器官である中心体近くの繰り返し配列(次回紹介する)が濃縮されていることだ。実際染色体上にNADをマップすると、中心体と接した領域にNADが存在するのがわかる。このように、NADもランダムに決められているのではなく、どの領域が核小体とコンタクトするのかも一定のルールで制御されていることが理解できる。ヒトHeLa細胞のNAD内には1300近くの遺伝子があるが、NAD内にある遺伝子の全般的傾向としては、LADと同じで、遺伝子密度の低い領域が濃縮されていると言っていい。またNADに結合しているヒストンの修飾を調べると、H3K4me3やH3K27me3などの抑制性のヒストンが濃縮されており、LADと同じようにこの領域の遺伝子発現は抑制的に支配されているといえる。NADに存在する遺伝子で最も特徴的なのは嗅覚遺伝子だが、ほとんどの細胞で発現が抑制されている嗅覚受容体遺伝子が濃縮されていることも、NADが遺伝子を抑制するための構造化の一つであることを示唆している。一方、ヘテロクロマチン構造により抑制されているトランスポゾンは、特に領域を限らず均一に分離している。NADも研究が始まったばかりで、今説明した現象の分子メカニズムはほとんど分かっておらず、今後の研究が必要だ。
3回にわたって、遺伝子の配置、ヌクレオソーム、TAD, LAD,NAD,そしてクロモゾームテリトリーと、ゲノムが様々なレベルで構造化されており、構造化されることで、おなじ塩基配列でも転写される情報とは違った情報を生み出すことも見てきた。また、LAD, NAD,そしてクロモゾームテリトリーのように核の構造との相互作用により決まるケースでは、部分が全体を構成し、全体が部分を組織化するという生命特有の性質を見ることができる。この生命特有の部分と全体、原因と結果問題は、進化過程を通してしか説明がつかないが、幸いそれぞれの構造は進化の異なる段階で誕生したことが、これら構造の系統学からわかる。この意味で、ゲノムを様々なレベルで構造化し、新しい情報を開拓してきた進化過程の研究は重要分野になる気がする。
[ 西川 伸一 ]
構造部材としてのセントロメアDNA
2015年8月3日
ここまでゲノムには、転写して利用される情報だけでなく、構造(あるいは順序、位置など)としての情報も含まれていることを示してきた。今回は構造としての情報を考える最終回として、例えば家の柱や壁のような構造部材としてDNAが使われている場合について見てみよう。
すでにゲノムの一部はヘテロクロマチンという構造を取り、他の部分と比べてコンパクトに折りたたまれ転写が抑えられていることを紹介した。ただゲノムの中には常にヘテロクロマチン状態をとる領域と、細胞や分化段階に応じてヘテロクロマチン構造から解放され、転写が可能になる領域の2種類が存在する。常にヘテロクロマチン構造をとっているゲノム部位を調べていくと、全く遺伝子が存在しない大きな領域が見つかる。この領域のうち代表的なものが、セントロメアとテロメアで、共に真核生物の細胞分裂時に重要な働きをする。ここでは、セントロメアに焦点を当てて見てみよう。
まずセントロメアとは何か。真核細胞はDNAの倍加後、2つの細胞に分裂する。その時、細胞の中心に並んだ染色体を両方の細胞へと正確に分配する(図1)。
図1 細胞分裂中期。中央に並んだ染色体に微小管が結合して染色体を分離しようとしている。緑色蛍光は微小管、青色蛍光は染色体、赤い蛍光は動原体を染めてある。動原体は染色体セントロメアと微小管を結合させる構造である。(Wiki Commonsより)
この時、2つの細胞に別れた中心体から放射する紡錘糸(微小管)はキネトコア(動原体)を介して染色体と結合し、細胞分裂の経過とともに染色体を両方の細胞へと引っ張り込む。この動原体と染色体の結合の中心になっているのがセントロメアと呼ばれる場所のヌクレオソームだ。他の部位のヌクレオソームと比べると、セントロメアのヌクレオソームにはcenH3と名付けられた特殊なヒストンが存在している。このcenH3は動原体を構成する分子と結合して染色体と紡錘糸を結びつける役割を持っている。cenH3が普通のH3と置き換わって存在しヘテロクロマチン構造をセントロメアと呼んでいる(図2)。
図2 セントロメアの構造。セントロメアではヒストンH3の一部がcenH3に置き換わっている(この図は私が徹底的に単純化しており、実際には様々な割合でcenH3が置き換わっている)。このcenH3に動原体を構成する多くの分子が結合し、微小管と染色体を結合している。
私たちヒトの細胞ではセントロメアの数は染色体に一カ所あるだけだが、この様式は全ての生物に共通というわけではなく、例えば線虫では一つの染色体上に何カ所も形成される(図3)
図3 線虫とヒト染色体のセントロメアの位置をcenH3染色で示している。ヒトでは染色体中央に一箇所のセントロメアがあるが、線虫では一本の染色体に複数のセントロメアが存在する。
動原体は分裂が終わると消失する。したがって、分裂ごとに同じ場所にセントロメアが再構成される必要がある。即ち、cenH3とヘテロクロマチン型ヒストンが特定の領域のDNAと結合し特殊なヌクレオソームを構成する必要がある。この決まった場所に繰り返しセントロメアが形成されるメカニズムは完全にわかっているわけではない。最初、酵母を用いた研究では、セントロメアには一定の塩基配列が存在することが示され、セントロメア型ヌクレオソームは特定の塩基配列にガイドされて形成されると考えられた。しかし、ゲノムプロジェクトが進むことでセントロメアに特有の遺伝子配列があるという考えは揺らぐ。例えば、ヒトのセントロメアには171bpの長さのαサテライト配列と呼ばれる配列が繰り返しており、最も長い場合で5Mbにわたって存在している。マウスでも同じようなサテライト配列の繰り返し構造が存在することから、サテライト配列の繰り返し構造がセントロメア型ヌクレオソームをガイドする構造であると考えられた。しかし、馬の染色体の中にはこのような繰り返し配列が全く存在しない染色体が存在する。cenH3をリクルートするゲノム側の条件を理解することは一筋縄ではいかないことがわかった。いずれにせよ、セントロメアやテロメアはそれ自体何かをコードしているわけでもなく、また周りの遺伝子を構造化しているわけでもない。セントロメア領域から、様々な蛋白分子と共同して純粋に染色体の構造部材として働いているゲノム部分があることを理解していただいたのではないだろうか。
さて、染色体の構造部材としてのセントロメアはペリセントロメアと呼ばれるやはりヘテロクロマチン構造をもつ領域に挟まれている。ペリセントロメアは染色体が分離する際、染色体同士を結合しているコヒーシンと呼ばれる分子が最後まで残って結合している場所だ。このおかげで染色体が娘細胞に分配される時、最後に動原体部分が外れ、完全な染色体を分配することができている。この意味で、ゲノムのペリセントロメア領域もゲノムを支える構造部材と考えることができる。では、ゲノムのペリセントロメア領域をヘテロクロマチン化するための条件とはなんだろう?興味深いことに、セントロメアやテロメアと異なり、ペリセントロメアにはLINE,SINEを含む様々な繰り返し配列とともに、染色体の他の場所から転座してきた普通の遺伝子領域も見つかっている。したがって、繰り返し配列が存在するからヘテロクロマチン構造が生まれるのではなく、この領域自体が飛び込んできた遺伝子をヘテロクロマチン構造に変換する力があることを示している。このメカニズムについてはゲノムとRNAの関係という枠組みで次回に考えるが、飛び込んだ遺伝子をヘテロクロマチン化することで構造部分を拡大できる巧妙な仕組みが可能になっている。この観点に立つと、私たちゲノムの半分を占めるトランスポゾンの一部は構造部材として使われている可能性もある。というのも、ほとんどのトランスポゾンにヘテロクロマチン構造を取らせる仕組みを私たちは有している。この意味で(これは私の戯言だが)トランスポゾンは構造部材を新たに調達するにはうってつけの材料に思える。
3回にわたってゲノムが構造化されること自体新しい情報を生むことを見てきた。構造部材としてのDNAまでが情報と呼べるのは、構造も、材料も、コードも全て同じDNAを媒体とする線状の情報に置き換え、子孫に伝えるからだ。最初ただの物質でしかなかったDNAが様々なモノやコトと対応できる情報に転換する。最初DNAがコードとして何と対応付けられていたのかは21世紀重要な問題だ。これは私の戯言だが、おそらく最初に対応付けられたのはアミノ酸の並びではないような気がする。というのも、この対応関係はRNA,tRNAを必要として、あまりにも複雑だ。とすると、今セントロメアでみられるような部材としてのDNAと蛋白の関係と、DNA,RNAの関係が最初は別々に存在していたと考えたほうがわかりやすい。次回はこの問題を横目で睨みながら、ヘテロクロマチン転換のメカニズムから初めて、DNAとRNAの関係について考えてみたい。
[ 西川 伸一 ]
DNAとRNA
2015年8月17日
DNAとRNAと聞くとすぐセントラルドグマ、すなわちDNA〜RNA〜ペプチド(タンパク)と続く情報から実体への流れを想像する。しかし、太古の昔、地球上で核酸が情報の媒体として使われ始めたとき、セントラルドグマに定式化されているDNA,RNA, タンパクの間の美しい関係があったとは考えにくい。DNAは地球上に物質として生まれた時点から、複製によって正確に伝えるという点ではすでに情報媒体としての条件を満たしている。ただ、最初から塩基配列のコードとアミノ酸が対応していたとは、神でも想定しない限りまず信じられない。すでに見てきたように、DNAはタンパク質のコード以外にも様々な情報を表現し、それを安定に子孫に伝えることができる。言い換えるとDNAはこれらゲノムの持つ全ての機能を最初に複製可能にした物質だったように思える。DNAが作り出す構造と、タンパクやRNAが、コードとは無関係に相互作用できることは現存のヌクレオソームを見れば明らかだ。一方、RNAとなるとさらに多様な構造を取り得ることがわかっている。実際RNAだけから特定の酵素活性を持つ構造を作ることが可能で、最初の生命体はRNAでできていたと考える人もいる。ただ生命がRNAから始まったという主張に完全に同意できないのは、複製という点でRNAが様々な問題を抱えているからだ。私自身は、まず物理化学法則に従って太古の昔にDNA,RNA,タンパク質が独立してランダムに生まれ、それが何か関わりあい続けるうちに突然情報体系への大転換を遂げたように思える。最近進んだノンコーディングRNA(ncRNA)の研究から、ncRNAがDNAやタンパクと直接関わりあって染色体構造を変化させているのを知ると、この過程に太古の3者の関係の名残が反映されているのではと妄想を巡らせてしまう。そこで、最後にこの妄想に付き合ってもらうためにncRNAが誘導するヘテロクロマチン化について見ておこう。
細胞が発現する全てのRNAを特定しようと試みたFANTOMやENCODEプロジェクトが終わって、全ゲノムの8割にも及ぶ領域が転写されているという事実が発表された時、私たちは何かの間違いではないかと驚いた。しかし前回紹介した、ヘテロクロマチン構造により固く閉ざされたゲノム領域であるセントロメアやペリセントロメア領域ですら転写されていることがわかり、低いレベルなら、クロマチンの構造とは無関係にゲノムのほとんどが転写されていることが認められるようになっている。ヒトセントロメアには174bの繰り返し配列からなるマイナーサテライト配列が、サテライトII,IIIを含む大きなメージャーサテライト配列からなるペリセントロメアに挟まれているが、この二つのサテライト配列もノンコーディングRNA(ncRNA)として、転写されている(図1)。
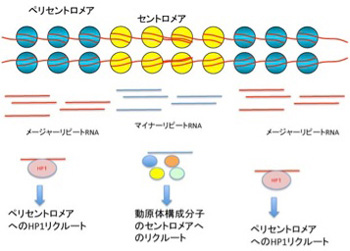
図1 セントロメア、ペリセントロメアからncRNAが転写され、それが自らのヘテロクロマチン構造形成をガイドする。
これまでの研究で、このncRNAがセントロメア、ペリセントロメアのヘテロクロマチン構造に重要であることが明らかになっている。例えば、マイナーサテライト配列から転写されるRNAはセントロメアにcenH3をリクルートするとともに、CENP-Cを含む様々な動原体の構成分子をこのヌクレオソームに結合させる機能を担っている。すなわち、セントロメアからRNAが転写されないと、正常のセントロメア・動原体の形成ができない。同じように、ペリセントロメアのメージャーサテライト配列から転写されたncRNAは、ヘテロクロマチン転換を誘導するタンパクHP1(ヘテロクロマチンプロテイン1)をペリセントロメアにリクルートし、ヘテロクロマチン構造を形成する。このようにそれぞれの領域の特異的なクロマチン構造を形成し維持するためには、低いレベルでRNAが転写されていることが必要だ。では、クロマチン構造を調節しているのは必ずその領域のゲノムから転写されるRNAだろうか?答えはNoで、様々なRNAがヘテロクロマチン誘導に使われていることがわかっている。最も大掛かりで典型的な例がX染色体不活化(XCI)なので次に見てみよう。
ゲノムからあまり間違うことなく予想できることの筆頭は男女の区別だ。男性はXY、女性はXXが人間の性染色体で、この差が男女を決めている。ただこの方式には一つ問題がある。すなわち男性にはXが1本に対して、女性は2本という数の差だ。というのもXには常染色体とおなじように生命維持に必須の遺伝子が存在するため、何もしないとX染色体上の遺伝子の発現量は女性が2倍になる。多くの遺伝子は発現に2倍の差があっても問題ないが、中には発現を男女一定に保っておく必要のある遺伝子もある。私たち哺乳動物はこの課題に、女性の片方のX染色体をすべて不活性化することで答えている。この不活化にはX染色体の不活化センターと呼ばれる部位に存在するXist遺伝子が主役を演じている。Xist遺伝子は8個のエクソンからなり、普通の遺伝子とおなじようにスプライシングを受けた後、polyAも普通に付加される(図2)。
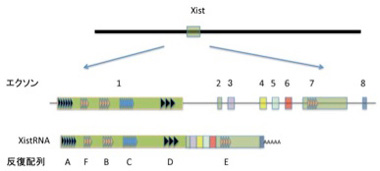
図2 X染色体のX染色体不活化センターに存在するXist領域は、発生過程で片側の染色体だけで転写が始まり、転写されたXistは普通の遺伝子と同じように8個のエクソンを持ち、転写後スプライシングされ、ポリAが添加されるが、核外には移動しない。
Xist RNAが他の遺伝子由来の RNAと異なる点は、RNAが核外に移動することはなく、従って翻訳もされない。転写が片方のX染色体だけに限られ、またもう一方のX染色体に結合しない巧妙な仕組みについて理解は進んでいるが、全て割愛して、とりあえずXist遺伝子から転写されたRNAは片方のX染色体全体を覆っていると理解してもらおう(図3)。
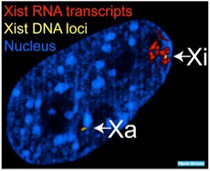
図3 メスの細胞では、片方の染色体だけがXistによりカバーされている。黄色で示されているのはXist領域で、両方の染色体に存在するが、XistRNAは染色体全体に広がっている。
(Wiki commonsより)
先に紹介したセントロメアや、ペリセントロメアはその場所から転写されたRNAがホスト領域と結合するが、このXistは17kbの領域からだけ転写されるncRNAが染色体全体に結合する。したがって、Xist-RNAはDNAと塩基配列の相補性で結合するのではなく、Xist-RNA内に存在する繰り返し配列Cによりヌクレオソーム自体に結合することがわかっている(図2)。また、X染色体を覆うといっても実際にXistは2000個ぐらいしか発現していないことがわかっており、おそらくX染色体に飛び飛びに結合して、その場所からヘテロクロマチン構造を拡大していくのだろうと想像されている。実際Xistには遺伝子のサイレンシングやヒストンのメチル化に関わる分子をリクルートする繰り返し配列が何種類も存在しており、図2に示したrepeatAは遺伝子のサイレンシング、repeat F, Bはヒストン修飾に関わる分子、そしてrepeat CがXistがクロマチンと結合するのを媒介することがわかっている。このように、DNAとタンパク、ncRNAとタンパクがそれぞれ複合体を形成し、さらにこの両者が働き合うことで、ヘテロクロマチン構造が形成されていくのがわかる。この3者の相互作用には塩基配列コードは必要ない。
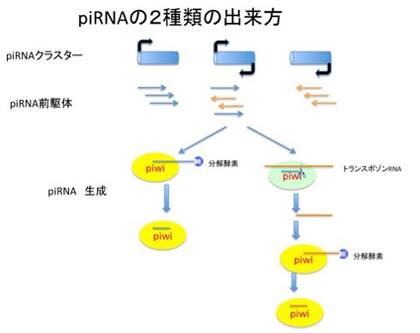
図4PiwiRNAの形成と機能
私たちのゲノムの5割はトランスポゾンで占められているが、最後にゲノムに飛び込んできたトランスポゾンをヘテロクロマチン化するメカニズムを担うPIWIタンパクとpiRNAについて見てみよう。海綿から人間まですべての動物に存在して、植物には存在しない遺伝子の一つがPIWI分子で、トランスポゾンの活動を抑える細胞レベルの防御反応の主役になっている。この時に働くRNAがPiwi interacting RNA (piRNA)で、のちに述べるようにこれは遺伝子の機能を抑制するための様々な分子を標的DNAやRNAにガイドする役目がある。このpiRNAは主に生殖細胞に発現している25-26のヌクレオチドからなる小さなRNAで、その種類は千種以上存在すると考えられ、2種類の方法で作られる(図4)。最初の方法は、ゲノム中に存在する200−数百kbのpiRNAクラスターと呼ばれる領域から転写される長いRNAがPiwi分子と結合して、短い機能的な長さに切り整えられることで作られる。このpiRNAクラスターには様々なトランスポゾンに実際に存在する領域が集められており、一種のトランスポゾンのカタログのような領域だ。従ってこのカタログ領域から転写されPiwi分子により処理されてできるpiRNAはトランスポゾン領域と相補的に結合することができる。このカタログのおかげで、ほとんどのトランスポゾンの配列を認識して、遺伝子発現を抑えるための機能分子をその領域へガイドすることができる。piRNAは、このクラスター外のトランスポゾンが活性化され転写が始まったRNAからも作ることができる。この時、トランスポゾン転写産物は、すでにその細胞に存在している相補的なpiRNAとpiwi分子上で会合し、トランスポゾンからの転写産物を短くカットする。こうして出来た短いRNAはPiwi分子上でさらに処理を受け、新しいpiRNAに変換される。どちらの方法でも、出来てきたpiRNAは様々なレトロトランスポゾンと相補的なガイドRNAとして働くことができる。
さてこうしてできたpiRNAとPiwi分子複合体は、piRNAによりガイドされてきた領域のヌクレオソームをヘテロクロマチン型に転換することができる。この場合、標的の情報はこれまで経験したトランスポゾン配列のライブラリーとしてpiRNAクラスターに記録してある。また、今活動しているトランスポゾン由来のRNAもpiRNAに変換してガイドに使うことで、トランスポゾンと他の領域を区別し、トランスポゾンだけをヘテロクロマチン化するのに関わっている。
以上の3つの例から、ncRNAが特定の遺伝子部位の染色体構造を決めるために重要な働きを演じているが、その形成方法と作用機序については多様性が大きいことがわかってもらえたのではないだろうか。さらにpiwiとpiRNAのペアには、ヘテロクロマチン化だけではなく、RNAによりガイドされて直接トランスポゾンのDNAをメチル化し、すでに転写されてしまったトランスポゾンRNAに結合してそれを分解する働きがあることがわかっている。この意味でpiRNAは、多様な武器をトランスポゾン攻撃に集中してその活動を抑える最前線の兵士のようだ。おそらく専門知識のある読者は、piwiとpiRNAの説明を見て、原核生物や古細菌に見られる外来遺伝子の侵入に対する防御機構、CRISPER/Cas9システムを思い出したのではないだろうか。次回は単細胞動物へとさかのぼってセントラルドグマとは異なるDNAとRNAの相互作用について調べてみよう。
[ 西川 伸一 ]
ゲノムの自己
2015年9月1日
塩基配列が何を意味しているかわからないという理由だけでジャンクと名付けていた領域の中で、機能が明らかになって大ブレークしたのがクリスパーだろう。最初大腸菌ゲノムの中の不思議な繰り返し配列として見つかった配列(まさに当時はジャンク配列)が、実際には大腸菌に感染する様々なウィルスやプラスミドの断片であることがわかり、クリスパー上流にあるCas遺伝子群とともに、外来DNAを自己のゲノムから区別して抑制する一種の免疫システムを形成していることが明らかになった(図1)。
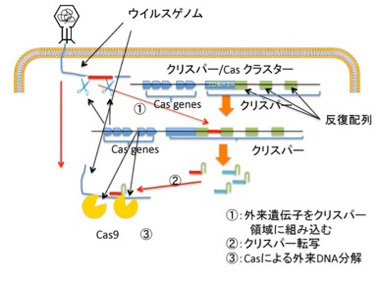
図1 外来遺伝子への免疫システムとしてのクリスパー/Casシステム
説明は本文参照。
外来の遺伝子が侵入すると、その一部を切りとって、クリスパー領域に組み込むことで、侵入記録を遺伝子ライブラリーとして記憶(図1①)、次の侵入に備えるという免疫の仕組みだ。こうして形成された外来遺伝子のライブラリーは、Cas9と結合するRNAリピート領域と一体となったガイドRNAとして転写され、次に相補的外来遺伝子が侵入してくると、その遺伝子とハイブリダイズしてCas9を侵入遺伝子へと導き(図1②)、これを切断、分解する(図1③)。この最後の反応を利用して開発されたのが、遺伝子改変技術を根本的に改革したゲノムの配列特異的編集法で、今や医学応用を含めあらゆる生命科学分野で大ブレークしている。
さて外来遺伝子をライブラリーとしてゲノムの中に保持し、そこから転写されたRNAを標的遺伝子の抑制に使うという本来のクリスパーシステムを理解すると、まさに前回紹介したPiwi/piRNAとよく似た系だとわかるはずだ。まずどちらも自己と他を区別するための記憶として、外来遺伝子配列を特定の領域に蓄積し、それを自己ゲノムと他を区別するのに使っている。これも最初は機能がわからずジャンク配列とされていた。この例から、他のジャンク配列もまだまだ思わぬ機能が明らかになる可能性がある。進化では、役に立たないと決めつける方がいつも間違っていることが多い。
さて、クリスパーやPiwi/piRNAの例から再認識刺されるのは、情報としての自己のゲノムは、他のDNA情報に常に開かれていることだ。いつでも他のゲノムは自分のゲノムの中に侵入してくる。このため常に自己の情報と他の情報を区別して、自己の情報だけを守る仕組みが生命誕生後の早い時期から存在している。驚くべきことに、この機構を拝借して逆にホストのゲノムを分解する能力を獲得したしたたかなウイルスすら存在している。このphageウイルスは幾つかのCas遺伝子と侵入したバクテリアが持つ防御最前線で戦う分子PLE遺伝子をアタックできるクリスパー配列を持っている。すなわち、本来バクテリアがウイルスゲノムの侵入を守るセットを使って、ウイルス抵抗性に関わる遺伝子を攻撃して自らを守っている。生命誕生は、自己と他という関係の誕生でもある。自己のゲノムを他から区別する仕組みはこのように、ウイルスを含むあらゆる生物へと拡大する宿命にある。
クリスパーからわかるように、ncRNA(non-coding RNA)はあらゆる生物で機能しており、生命誕生初期から、あるいはそれ以前からこのようなDNAとRNAの関係があったことをうかがわせる。しかし、なかでもゾウリムシやテトラヒメナのような繊毛虫類でのncRNAの大胆な使い方には驚かされる。最後に、テトラヒメでncRNAがどのように使われているか見てみよう。ただその前に、これまで紹介しなかったsiRNA(small interfering RNA)について、piRNAと比較しながら簡単に紹介しておく。Piwi分子の結合するpiRNAは、piRNAクラスターか活性化されたトランスポゾン由来だが、どちらも転写された一本鎖RNAだ。ただ、様々な方法でできてきた2本鎖RNAからも同じような遺伝子抑制性のRNAを形成することができ、siRNAとよばれている(図2)。
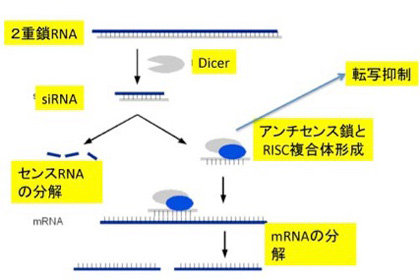
図2 siRNAの形成と作用:Wikiコモンズから改変
この系では、2本鎖RNAはDicerと呼ばれる分子で決まった長さに切断された後、piwi分子の代わりに、RISCと呼ばれる分子にロードされる。このアンチセンスRNAとRISCの複合体は、このRNAをガイドとして、外来RNAの分解や、標的遺伝子のヘテロクロマチン化に関わっている。ガイドに利用するRNAが最初2本鎖である点がpiRNAと異なるが、共にトランスポゾンの活動を抑制するメカニズムだ。この2つのメカニズムを頭に入れておいて、次に繊毛虫類の一つ、テトラヒメナを例に驚くべき精巧なncRNAの利用法を見てみよう。これから述べる話のほとんどはウィーン・IMPに在籍するこの分野の第一人者、望月一史さんたちの総説を参考にしたことを断っておく。
さてゾウリムシやテトラヒメナは、一つの単細胞の中に大核と小核の2種類の核を持っており転写は大核からだけに限ることで、高い転写効率を達成している。以後は望月さんたちの研究しているテトラヒメナに限って見ていこう(図3)。
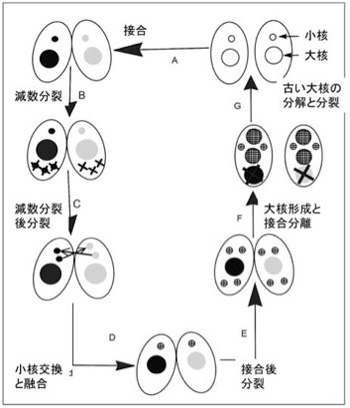
図3 テトラヒメナの接合による性生殖:説明は本文、Wikiコモンズ改変
さて栄養が豊富な環境では、テトラヒメナは分裂により増殖する。この時、大核も小核も同じように複製され、両方とも娘細胞に分配される。一方、栄養条件が悪くなると、2つのテトラヒメナが接合し二つの個体で遺伝情報の交換を行う。おそらく異なる個体のゲノムを合わせることで、厳しい環境に適応しやすくするための戦略と考えられる。しかしこの時、高等動物の減数分裂に当たる過程が小核だけで起こる。Wiki commonsからの図を参考にして接合過程を見てみよう。まず図の左上から、減数分裂により一倍体の小核を最終的に1個形成する(残りの小核は全て分解される)。次に小核が分裂して一つの細胞の中に2個の1倍体の小核ができる。この間、大核はそのままで転写を続け細胞を維持している。次に、それぞれの個体は小核を交換し合って、最終的に別々の個体由来の一倍体の小核が融合した2倍体の小核が生まれる。この小核はその後2回の分裂により4つの小核を作り、そのうち2つが新しい大核を形成するのに使われる。この小核から大核への転換時に、それまで転写をまかなっていた大核は分解する。こうして生まれた新しい2つの小核と2つの大核を持った個体が次に分裂することで、接合した結果できた新しい遺伝情報を持った一個の大核と一個の小核を持つ2つの個体が生まれるという複雑な過程だ。
なぜこれほど複雑な接合過程を持っているのか不思議だが、これは大核のゲノムと小核のゲノムが異なっていることに起因する。正確に言うと、小核中のゲノムには全域にわたって6000以上の0.5-30kbの遺伝子断片が挿入されていて、IES(internal eliminated sequence)とよばれている。IESもクリスパー・piRNAと同様にトランスポゾンや繰り返し配列から出来ており、これまでの定義ではジャンクと呼ばれる配列だ。しかし、IESは有性生殖の初期以外の時期において、小核からの転写を抑制する積極的役割を持っている。すなわち全てのジャンクはジャンクであることで機能を持つ。一方、小核から大核ができるとき、新しい大核内の全てのIES(=ジャンク)が正確に除去されないと、遺伝子の転写ができない。実際、大核にIESが残ってしまうとテトラヒメナが死んでしまうことがわかっている。これに加えて、大核のゲノムは250ぐらいの断片に分解され、それぞれは増幅されているおかげで、転写レベルを上げるよう改変されているが、この過程についてはここでは深追いしない。
ではどのようにしてIESは小核ゲノムから正確に除去されるのだろうか?ここにDicerやPiwiに似た分子と、小核から転写されるncRNAが一役買っている(図4)。まず新しい大核への転換が始まる前に小核全体が双方向性に転写され、ゲノム全域をカバーする2本鎖RNAが形成される。この2本鎖RNAはDicerに似た分子Dcl1pによって小さく断片化されncRNAになり細胞質へと移行し、piwiに相同な分子Twi1pに結合する。Twi1p上でガイドRNAだけを残してパッセンジャー鎖は除去される。こうして形成したガイドRNAを結合したTwi1pは大核に移行し、大核中のゲノムと比較され、大核中に存在する配列が全て除去される。この結果残るのはIESに対応するRNAをと結合したTwi1pだけになる。
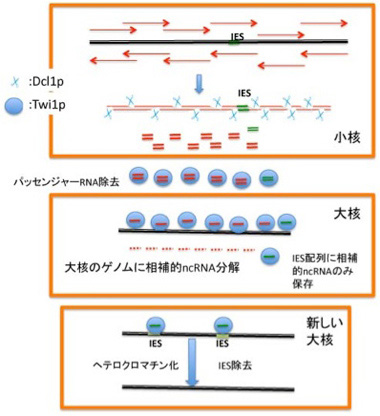
図4ncRNAを使った新しい大核形成時のIES除去機構:説明は本文
この選択が実際にどう起こっているのかは研究途上にあるようだが、大核のゲノムと相補的なRNAはゲノムDNAによりTwi1pから引き剥がされ、分解されると考えられている。メカニズムはともかく、これにより大核ゲノムと小核ゲノム間で引き算を行うことができ、結果として全てのIESに相当するncRNA をロードしたTwi1pが残る。次にこのTwi1p/ncRNAは新しく形成された大核に移行して、今度はIESに結合し、IESをヘテロクロマチン化する(H3K9, H3K27のメチル化)。このヘテロクロマチン化が標識となって、IES配列がゲノムから全て除去される。残念ながら、ヘテロクロマチン化された領域がどのようにゲノムから切り離されるかについての具体的なメカニズムについてはまだまだ明らかでない。しかし、テトラヒメナでは小核ゲノムからIESを大規模かつ正確に取り除く目的でncRNAが利用されることがわかる。
IESの多くがトランスポゾンを起源としていることを知ると、テトラヒメナではトランスポゾンが自己ゲノムを調節する機能ユニットとして積極的に活用されていることがわかる。テトラヒメナの2核構成も情報論から見ると面白い。一般的に情報理論は、伝えるためのコミュニケーション理論とオペレーションのためのコンピュータ理論に分かれる。ゲノムもそうで、子孫に伝える役割と、細胞のオペレーションとに役割を演じ分けている。問題はオペレーションのための情報は、伝えなければならない情報のほんの一部だということだ。そのため、真核生物はエピジェネティックな機構を進化させ、様々な方法でオペレーションに必要でないゲノム領域をオペレーションから隔離し、オペレーションの邪魔をしないよう抑制している。しかし、分裂のたびにエピジェネティック標識を正確に再構築するためには複雑な機構が必要になる。その代わりに、必要のない領域を全て除去した大核と小核に分けて、オペレーションと伝達を完全に分離してしまう戦略は、一見複雑に見えても、実は最も簡単で安心な戦略なのかもしれない。実際、テトラヒメナは普通、両方の核をただ単純に複製し続ける。そのとき、エピジェネティック状態を常に再構築しないで済むことの利点は計り知れないはずだ。このように情報のコミュニケーションとオペレーションを分離したテトラヒメナは驚くべき生物と言えるだろう。
これでゲノムの構造については終わり、次からはゲノムの働きについて考えていく。
[ 西川 伸一 ]
ゲノムの働き
2015年9月15日
ゲノムの解剖学に続いて、次はその働き方を見てみよう。前回紹介した、ゲノム情報を大核と小核に分けて使っているテトラヒメナは、ゲノムの働きを理解するための優れたモデル生物になっているので、これを参考にしながら見ていこう。
ゲノムの働きの第一は、複製を通して親と同じゲノム情報を子孫に伝えることだ。無生物と比べる時、生命が示す特徴の一つは複製(replication)だ。私たちの細胞のように有糸分裂で増殖する細胞の場合、細胞分裂時にほぼ正確に複製され、子孫細胞に分配されるものに、ゲノムと中心体がある。しかし、原核生物になると複製分配されるのはゲノムだけになる。生物全体を通して見たとき、複製されて子孫に伝達されるものは結局ゲノムしかない。このことから、ゲノムの働きは親と同じ状態を増殖させるためコピーして子孫に伝える情報としての働きがあることがわかる。
しかし正直に言うと、なぜ生物は分裂し、増殖しようとするのかについてはよくわからない。熱力学に逆らって、平衡に陥らないようにメタボリズムを維持するだけなら増殖は必要ない。実際、全く分裂しない赤血球は200日の寿命があるし、神経細胞のように数十年にわたって分裂なしに生き、新陳代謝も盛んな細胞は存在する。ただ、複製のない細胞システムはどれほど新陳代謝が維持できたとしても、環境が変化してしまえば物理的に滅びる。環境に適応し、さらには環境まで変えてしまえる可能性があるという点で、結局は複製能力を持つシステムだけが地球上で生物として存続できたのだろう。
一方複製自体に情報の複製は必要ない。例えばウイルスの被膜を考えてみよう。被膜形成に必要な幾つかの構成タンパクが必要なだけ調達できるなら、一定の大きさのウイルス被膜は自然に形成される。ここでもし被膜が物理的強度の問題で一定の大きさになると壊れると仮定しよう。不完全な被膜は、壊れた一部を核にしてもう一度完全な新しい被膜を形成できるだろう。すなわち、平衡から遠く離れた条件さえ満たせれば、形成、破壊、再形成を繰り返して増殖するウイルス被膜のような構造は十分考えることができる。同じように、例えば核のないプロトセルのような液胞が一定の大きさになると物理的にちぎれるとしよう。このちぎれる限界の細胞内成分の量と膜の強度(この時膜も自然に成長するとしておく)が物理的に決められておれば、情報はなくても一定サイズになると2個の液胞が作られ、またその液胞が分裂するというサイクル、すなわち複製サイクルを考えることは可能だ(実際そのような実験が行われている)。しかし実際には、何らかの情報がないと同じものを作り続けることは難しく、生成されるプロトセルの大きさも時間とともに大きくばらつくだろう。判を押したように同じ大きさのプロトセルを作り続けるためには、デザインや機能を指示できる情報が増殖には必要になる。
もちろん情報といっても、最初から核酸が情報媒体を担っていたと考える必要はない。例えばたんぱく質の構造を変化させるだけなら、プリオンのような例もある。しかし、増殖・生存に必要な情報を、核酸媒体を使って細胞の化学的オペレーションから完全に分けることに成功した私たちの先祖だけが結局地球上で生き残れた。この結果、現存する全ての自立生命は、DNAをゲノム情報の媒体として使っている。このように、ゲノムの働きの第一は、生物の増殖を支えることになる。
テトラヒメナにもどろう。テトラヒメナは単細胞生物で、通常は分裂により増殖する。このとき、大核も小核も同じように複製され、それぞれの娘細胞に分配される。では両方の核が複製されることはテトラヒメナの維持に必要なのだろうか。例えば、小核を除去したテトラヒメナは増殖が可能だろうか? 実験が行われたかどうか明らかではないが、おそらくかなりの期間分裂を続けることはできるだろう(原理的には無限に分裂していい)。実際、核を一つしか持たない真核生物の方がこの世には多く存在し、増殖を繰り返している。テトラヒメナの場合大核ではゲノムが200以上に断片化されているなど、特殊な構造を持っているため、大核は長い期間にわたって安定な複製を維持しにくいかもしれないが、基本的には小核がなくとも増殖は可能だろう。
では小核は何をしているのか?前回説明したように、小核は二つの個体の接合時に独自の活動を始める。まず減数分裂を行い1倍体の核を2個形成する。次に、同じように減数分裂によりできた他の個体からの1倍体の小核を交換し合い、最終的に融合する。この結果、異なる個体のゲノムを集めた2倍体の核が作られる。この小核からまず新しい大核と小核を作り直したあと、その後は通常の分裂増殖サイクルに戻る。
この複雑な過程から出来るのは2倍体の小核なのに、なぜこれほど面倒臭いことをしなければならないのか。テトラヒメナの小核の場合、新たに何千箇所もの挿入配列を取り除いて大核まで作らなければならない。なぜこんな複雑な仕組みが選ばれたのか?一つの可能性については前回述べた。ここまで複雑な接合の仕組みが維持されていることは、テトラヒメナにとって最も効率のいい方法なのだろう。
もともと異なる個体間でゲノム情報を交換し合い、ゲノムレベルの雑種を作ることは種の生存のために重要だ。もちろん逆もあるだろうが、一般的に多様な情報を持っている方が多様な環境に対応できる。もう一つ重要なのは、これにより有害な突然変異を元に戻したり、薄めたりできる。ゲノムでは接合だけが異なる個体の情報を一つの個体に統合することを可能にしている。接合だけではそれぞれのゲノムは独立したままで、いわば2人3脚ではあっても本当の融合ではない。しかし、次の減数分裂が起こるときには、染色体間で相同組み替えがおこる。すなわち染色体同士で部分の交換が行われ、異なるゲノム由来の情報が一つのゲノムに文字通り融合する。テトラヒメナでは、このゲノムの統合、交換、融合を通したゲノム情報の間のコミュニケーションを小核が専門的に行っている(前回の図参照)。一方、ほとんどの真核生物は、これを一つの核で行っている。
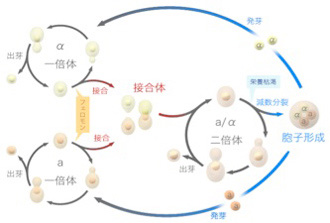
図1出芽酵母の減数分裂と接合。説明は本文(Wikicommonsより)
生物学ではこのゲノム間のコミュニケーションを「性」と定義している。ほとんどの真核生物ゲノムが共通に持つ働きだ(例外については最後に紹介する)。テトラヒメナの場合、個体全体が接合したままコミュニケーションが行われるが、より一般的な戦略は、コミュニケーション(接合)用の細胞が特別に分化し、その細胞同士で融合が起こる。またこの細胞を作る過程で減数分裂が起こり、ゲノムの交換と融合が可能になっている。例えば、出芽酵母では2倍体の分裂個体が減数分裂で1倍体の胞子を作り、この胞子が接合して2倍体の個体を作る(図1)。一方、生殖細胞が体細胞から分離した高等動物では、この減数分裂と接合は、生殖細胞系列の配偶子、例えば精子と卵子、植物では花粉や雌しべの役目になる。それぞれが分化し接合する方法は全く異なるが、全てで共通に行われているのは個体間のゲノム情報の交換だ。面白いのは分裂個体が減数分裂サイクルに入るのはほとんどの場合、外界の栄養状況が悪くなるなど逆境に晒された時だ。すなわち、逆境では他の個体と情報交換することで、自分にはない能力を取り込める可能性がある。この異なる個体間でゲノム情報を交換するコミュニケーション能力が、生存に負の作用を持つ突然変異を薄め、ゲノムの多様性を増大させてきた。まとめると、ゲノムの働きの第2は個体間でゲノム情報を交換することだ。
ゲノムの働きの第3は多様化して進化の原動力になることだが、この話題は次回に回す。今回は、2番目の働き、性、すなわちゲノム間のコミュニケーションを全く行わない生物のゲノムについて最後に紹介して終わる。2年前の8月Natureにワムシの一種belloid rotifer(Adineta vaga)ゲノムについての論文が掲載され、注目を浴びた(Nature, 500:453-457, 2013, 幸いこの論文はcommonsとして図の掲載が許されている)。注目を浴びた最大の理由は、このワムシに性別や、雌雄同体や、減数分裂の気配が全くないからだ。すなわち、異なるゲノム間の情報交換なしにワムシが何千万年も生きてこられた秘密がゲノムからわかると期待したからだ。期待通り、ワムシは2重人格とも言えるゲノム構造を持っていた。
図2 ワムシゲノムに見られる対立関係にある遺伝子とオオノログ(ohnolog)な関係にある遺伝子のマップ。ワムシの遺伝子の多くは、相同性の高いペア遺伝子(青色で示す)、すでに多様化が進んだペア遺伝子の2種類を持っている。
Nature, 500:453-457, 2013より。
図2には、この論文の図2bをそのまま掲載したものだが、ここで青い線で結ばれているのが相同性の強い対立遺伝子に相当する遺伝子ペアで、一方オレンジの線で結ばれているのが故大野乾博士の名前に由来するOhnologous遺伝子ペアだ。Ohnologous遺伝子とは、全ゲノムの重複後にコピーされた遺伝子を失わずに保持している重複遺伝子で、独立に進化し多様化してきた。ここでavとあるのはワムシゲノムでアッセンブリが確定している領域と考えて貰えばいい。重要なことは、ワムシでは対立遺伝子といっても二倍体のゲノムが存在するわけではなく、一つのゲノムの中に対立遺伝子セットが組み込まれていることだ。さらに、ゲノム重複によるペアも独立に存在するという不思議な構造をとっている。ゲノムは情報交換なしに単独で分裂を続けると、変異が蓄積して維持が難しくなる。ゲノム間で情報を交換しあうことは、この問題の解決のために必要で、真核生物は性生殖を通してこれを行う。性のないワムシでも、やはり情報交換が必要で、対立遺伝子の代わりに、重複した遺伝子を同じゲノム上で維持し、その間で情報交換することで異常が蓄積しないようにしている(図3)。
図3 有性生殖による異なるゲノム間情報交換と、分裂とゲノム内遺伝子転換による情報交換。Nature,
500:453-457, 2013,を改変
図3で示すように、この同一ゲノム内の相同遺伝子間の情報交換は、相同組み替えではなく、遺伝子転換で行われていると考えられている。相同遺伝子が同じゲノム上に散らばるワムシのようなゲノム構造が一旦できると、減数分裂、接合過程を介した情報交換はうまくいかないと考えられる。なぜなら、異なる個体の配偶子同士が接合したとしても、相同組み替えのための染色体のペアリングが正確に行えない。
この図からわかるように、同じゲノム上に存在していても、独立した対立遺伝子のペアを持つことで、ペアどうして相互に情報を交換して、有害な突然変異などを除去することが出来る。他にも、ゲノム内対立関係にある遺伝子と、それに対応するOhnologous遺伝子を比べると、相同性に大きな違いがあることから、相同組み替えがなくとも、遺伝子転換(gene conversion)メカニズムにより十分な多様性が得られることがわかる。
性生殖をやめたのが先か、ゲノム構造が特殊化したのが先かよくわからないが、結果的にワムシは有性生殖に頼らず、分裂に伴う遺伝子転換などの情報交換のみで多様化し、何千万年も生きることができている。面白おかしく考えると、ワムシは、普通なら異なる個体間に存在する対立遺伝子が一つのゲノムの中に統合されている2重人格者と言えるかもしれない。ただ、異なる個体間の情報交換をやめてワムシが何千万年も生きているからといって、性を通した個体間での情報交換が不必要かどうかはわからない。というのもワムシは普通の生物にはない不思議な性質をいくつも持っている。例えば、完全に乾燥しても水に触れれば生き返る。また放射線にも驚くべき抵抗性を示す。これほどの環境に対する耐性を獲得してしまえば、個体間の経験の差など取るに足らないのかもしれない。
[ 西川 伸一 ]
2015年10月1日
昨年5月、第6話で紹介したようにダーウィンの進化論の柱の一つは、集団内で形質の多様化が自然に起こるとする考えだ。ゲノムやDNAはおろか、遺伝の法則についても何もわからなかった時代にダーウィンがこれを着想したことに驚く。一方自然選択の概念は、マルサスや、適者生存による社会進化を唱えたスペンサーの著作からわかるように、当時の社会的思想風潮の一つではなかったかと思う。ダーウィンも適者生存の法則を説明するより、自然に起こる形質の多様化についての説得がはるかに難しいと予想していたのだろう。「種の起源」を集団中に自発的に起こる多様化の話から始めている。しかし、ゲノムの理解が進んだ今日では、自然に起こるゲノムの多様化(突然変異)が形質の多様化の背景にあることを疑うものは誰もいない。逆に自然選択については、適者生存といった単純なものではなく、形質の自然選択によるゲノム変化の研究は複雑で挑戦的な分野だと考えられるようになっている(後にできる範囲で議論したい)。進化がゲノムを持つ生物に内在する機能である以上、生物の多様化を推進する原動力がゲノムの3番目の働きになる。
私たちのゲノムに変異が導入される方法は、点突然変異から全ゲノム重複まで、極めて多様だ。具体的には、1塩基置換、欠損、挿入による点突然変異、小さな配列の挿入・欠損・逆位、染色体を含む大きな領域の欠失、挿入、転座、重複などがあり、全てがゲノムの多様化に寄与する。これほど多様な方法があるとわかっていたらダーウィンも苦労せずに済んだだろう。もちろんこのような変異は理論的可能性というだけではなく、実際に一定の確率で起こっていることは、多くのゲノムデータが蓄積されてきた現代では当たり前のことになった。ヒトゲノムを比べてみると、一塩基置換による特定部位多形性、すなわちSNP(single nucleotide polymorphism)は一千万箇所、小さな領域の挿入、欠損、逆転による変異は10万箇所に上ることが知られている。大きな染色体の変異となると、さすがにヒトゲノムデータベース内に見つけることはほとんどできないが、染色体重複だと例えば21番染色体の重複によるダウン症は有名だ。さらにがん細胞を調べると、様々な大きなスケールの変異を目にすることができる。例えば小児ガンの一つ神経芽腫の多くは13番染色体の特定部分が欠損することが知られているが、1983年に13番染色体全てが一本欠損した神経芽腫が発見されている(Science 219, 973-975, 1983)。他にも、ガンの遺伝的原因として染色体転座と呼ばれる異なる染色体同士の結合が知られている。例えば、ほぼ全ての慢性骨髄性白血病細胞は9番染色体と22番染色体の一部が結合した新しいキメラ染色体が形成される。これにより、bcr分子とabl分子が結合したキメラ分子が生まれ、これが血液幹細胞の増殖を促進し、白血病を引き起こす。こうして獲得される増殖優位性も適者生存の一つの形と言えるだろう。このような転座がガンの引き金になっている例は数え切れない。
ゲノムの多様化を考える時ガンゲノムから学べるもう一つの重要な事は、細胞が増殖を続ける中で驚くほど多くの変異が蓄積して行く事だ。例えば多数の悪性黒色腫の全ゲノム解読を行った論文によると(Cell vol161, 1681, 2015)、それぞれのガンごとに1Mbあたり平均16.8個の何らかの変異が見つかる。ヒトゲノムを3Gと考えると、この数字の3000倍、すなわち約5万個の突然変異が、ガンに蓄積していることになる。そしてこの中に異常増殖や転移性などに関わる機能的変異が複数個存在している。本当に個体の形質の多様化が目に見える形で起こっているだろうかというダーウィンの心配は全く杞憂でしかない。私たちのゲノムは猛烈な勢いで多様化するようできているようだ。
では、これほど様々な方法でゲノムを多様化させる原動力は何か?一時世間を騒がせたガン研究の大御所Vogelsteinたちが発表した論文にこのヒントがあるように思う(Science, 347,78, 2015)。私たちの体にはガンになりやすい組織となりにくい組織がある。この論文は、組織ごとのガンの起こりやすさと、その組織を維持するために必要な幹細胞の増殖回数をプロットして、両者が見事に相関することを示した論文だ。例えばガンの発症の多い直腸は、組織の維持のため幹細胞が10の13乗回近く分裂しなければならない。一方、小腸は長いように見えても幹細胞全体の全分裂回数は大腸の50分の1で、ガンの起こる頻度は100分の1であるのと一致する。すなわち、ガンにかかりやすさは、ガンが発症する組織の維持に必要な幹細胞の分裂数に比例し、つまるところ分裂時に起こる全く確率論的な変異により決まっているとする結論だ。もちろん生活習慣要因もガンのかかりやすさを変化させるが、背景となる分裂との相関は変わらないとした。これに対し、あまりに単純化しているとの非難の声が上がったが、私にとっては当然のことに思えた。というのも、ゲノムの複製は最も変異リスクが高い過程であることがわかっているからだ。2013年、7000にのぼる異なるガンのエクソーム解析からガンでどのような突然変異が起こっているか調べたNatureの論文(502,258,2013)でも、様々なガン共通に見られる突然変異の種類(例えばCからTへの変異)は、DNA修復や複製時に起こることが明らかになっている。DNA修復というと、私たちは放射線や紫外線、あるいは様々な化学物質などによるDNA損傷に対処するためのメカニズムだと思ってしまう。確かに地球上で暮らす限り損傷を避けることができないため、損傷を受けてもすぐに修復するための複雑な仕組みを開発してきた。実際、タバコや紫外線で修復が追いつかないほど損傷を受けるとガンの発生は上昇する。また修復酵素の力が弱まると少し紫外線に当だけでガンになる確率が高まる。その例が色素性乾皮症で、皮膚ガンの発症率は正常の20倍以上に達する。しかし普通の生活で起こる放射線照射や化学物質暴露よりはるかに強い突然変異誘発要因が存在している。それがDNA複製だ。詳しくは述べないが、DNA複製は2本鎖がチャックで開いたり閉ったりするように連続的に起こるものではなく、3種類のポリメラーゼ(一つはプライマーゼとも呼ばれている)が関わる複雑な不連続過程だ。この解明には当時名古屋大学の故岡崎博士の貢献が大きく、複製された短い不連続なDNA鎖を彼の名をとって岡崎フラグメントと呼んでいる(サイエンティストライブラリー岡崎恒子夫人の記事で詳しい解説有り)。最近DNAの変異が岡崎フラグメントの5’ 端に集中しており、これが複製間違いの多いポリメラーゼαで複製される短いフラグメントが完全に除去されず残ってしまうためであることが報告された(Nature 518,502,2015)。さらにポリメラーゼの複製間違いを修復する酵素(ミスマッチ修復酵素)の機能低下が多くのガンの引き金になっていることも分かっており、この酵素が先天的に欠損した小児のガンでは、普通のガンの何十倍もの突然変異が蓄積している(Nature Genetics 47,257, 2015)。これは点突然変異の話だが、染色体レベルの大きな変異の多くはやはり細胞分裂や減数分裂時に生じることがわかっている。これらの例から、形質の多様化の背景にあるゲノムの多様化を駆動するもっとも大きな原動力とは、生命に必須の細胞増殖そのものであることがわかる。言葉を変えると、複製されて子孫に分配されるというゲノムの働き自身が、多様化の原動力になっている。
最後に、生命の維持に必須のタンパク質や機能的RNAを作り続けるための情報としての働きがゲノムの4番目の働きで、テトラヒメナの大核が担っている。ただこの働きについては、ゲノムの解剖学で詳しく述べたので割愛する。
以上紹介したゲノムの4つの機能から見えてくるのが、ゲノムが時間スケールの異なる未来に向けての情報として用意されていることだ。まずゲノムは次の瞬間から未来に続く生命の維持のために、必要なタンパク質や機能RNAを必要な量だけ作り続けるための情報として働いている。次に、あらゆる生命は増殖を維持することが必要だが、ゲノムはこのとき様々な時間スケールで必要とされる細胞増殖の際に子孫細胞に分配するためコピーされる情報として働いている。以上二つのゲノム情報の働きが関わる時間はほぼオーバーラップしており、いずれも現在の環境に対応して生命を維持することに関わっている。ところが、テトラヒメナの小核の説明で述べたように、個体同士の情報交換の媒体としてゲノムは働いている。単細胞動物では、この交換は通常環境が変化したとき行われることから、もう少し長いスケールの時間で予想される環境変化に対応する情報として働いている。そして最後に、ゲノムは複製時に多様化する性質を身につけることで、遠い未来に備えるための進化可能性媒体として働いている。このようにゲノムは情報として様々な未来に備える働きをしている。面白いのは、転写や増殖のような近い未来に対しては、変化を抑えるよう安全弁を働かせ、一方、もっと長いスパンで予想される環境の変化に対しては自ら変化する性質を積極的に利用して備えている。
今回は、図のない説明になってしまったが、これでゲノムの働きについての説明は終わる。次回は、DNAを媒体とする生物の情報と、私たちが情報という言葉で一般的にイメージするものとの相違について見ていく。
[ 西川 伸一 ]

