2026年7月26日
自分のガンのRNAデータを見ていると、例えば分子Aは分子Bと結合するだろうかなどと疑問が出る。この分子間相互作用については膨大な研究があり、例えば K-ras がどの分子と結合するのかなどは文献を探すことでかなりわかるのだが、AとBをインプットして結合するかどうか判断してくれる AI があればさらに便利だ。
今日紹介する MIT からの論文は、まさにこのようなリクエストに正確に答えることができる AI モデルについての研究で、7月21日 Cell にオンライン掲載された。タイトルは「Expanding the scope of protein language modeling to protein-protein interactions with MSA Pairformer(タンパク質の言語モデリングをタンパク質同士の相互作用に拡大する MSA Pairformer )だ。
例によって数理部分をほとんど理解せず紹介していることを断っておく。
さて、ほとんどのタンパク質についての LLM は、個別のタンパク質を学習して、それらが分布する進化コンテクストの潜在空間を作っており、異なるタンパク質の相互作用に関するデータは存在しない。そのため構造を予測してから相互作用するか計算することになる。例えば KRAS と RAF1 が結合するかどうかを調べようと思うと、頭に浮かぶのは AlphaFold-multimer や AlphaFold3 を用いて構造を予測して相互作用の可能性を計算する方法だ。元になる AlphaFold は、構造を決定するとき、可能な限り多くの異なる種のアミノ酸配列を比較する Multiple sequence alignment (MSA) を作成し、またアミノ酸残基間の位置関係を記録する Pair representation を作成する。これから Transformer/attention で特徴抽出を行い、構造もデジュールで構造をアウトプットしている。従って、K-ras と Raf の構造を予測した後、結合面があるかを計算することになる。
この研究では、MSA と pair representation は一つのタンパク質の異なるアミノ酸残基の共進化と位置関係を表現しているので、わざわざ計算の大変な構造予測まで進まず、MSA と pair representation だけで、アミノ酸同士の結合を予測できるのではと考えた。さらに、可能な限り多くのアミノ酸配列の MSA ではなく、例えばヒトの K-ras に近いアミノ酸配列を重み付けして MSA や pair representation を形成することで、予測精度を挙げられると考えた。
そしてまず普通の LLM と同じように多くのタンパク質を学習させたあと、相互作用するか比べたいタンパク質AとBを、あたかも一本のタンパク質に統合して調べる、Paired MSA 方法を考案した。もしA、Bが相互作用している場合は、MSA を作成することで、セットで共進化するアミノ酸残基の組み合わせが見つかると予想され、見つかった場合はその残基同士が相互作用時の結合部位になっていると考えられる。もちろん全く共進化がないとすると、相互作用のないタンパク質ということになる。
この時、通常の Transformer と異なり、著者らが Query Biased Outer Product と呼ぶ方法を考案し、例えば調べたい human K-ras と Raf に近い配列で paired MSA を作成し、アミノ酸残基の関係が抽出された pair representationから相互作用するかどうか、どこで結合するのか、結合の強さはどうか等を計算することができるようにしている。
重要なのは、実際の構造予測をスキップしているおかげで、小さなワークステーションで済む点で、著者らは MSA pairformer では一つの H100 GPU で10日あればトレーニング可能で、大規模な ESM2 モデルのように512題の H100 GPU を使う必要がないことを強調している。
後は、既に知られている例えばバクテリアのトキシンとアンチトキシンや ABC トランスポーターなどいくつかのタンパク質について、分子間相互作用を数値化できるか、変異が起こったときの相互作用の変化を予測できるか、等を他のモデルと比較し、MSA pairfomer が高いパーフォーマンスを示すことを明らかにしているが、詳細は全て割愛する。
以上、数理については全く理解せずに紹介していることを繰り返すが、これまで一本のアミノ酸に限定されていた MSA や pair representation を、異なるタンパク質を一本に統合する Paired MSA を考案して、タンパク質同士の相互作用を正確に予測できるようにしたことは、私のような素人にもコロンブスの卵のような素晴らしいアイデアに思える。しかも、一般的な MSA のように可能な限り多くのアミノ酸のアラインメントをとるのではなく、知りたい (Query) 分子に近い (biased) タンパク質を重み付けしてアラインメントをとることで情報の質を高め、3次元構造生成をスキップして、計算コストを大幅に下げたということは、何でもかんでも大きなモデルを作ってそのパーフォーマンスを誇るのではない、新しい方向性を示しているように感じた。
おそらく私の説明では不十分だと思うので、是非自分で論文を読まれることを勧める。
2026年7月25日
ビクトリアの滝を案内してくれたザンビア人のガイドさんが、20種類の言語を話せると自慢していた。と言っても、英語以外はザンビアやジンバブエ周辺に住む人たちが使っている言葉のことで、それぞれはかなり似通っているおかげで何種類も話せるのだと思う。ろうあの子供から自然発生したニカラグアの手話は集団が200人ぐらいに達したときに誕生したと言われているが、ホモサピエンスは一定の集団になるとそれぞれの言葉を自然発生させるのだろう。とすると、膨大な数の言語が生まれ消えていったことになる。
今日紹介するハーバード大学とイエール大学からの論文は、完新世、即ち人類がユーラシアに分布した後2万年が経過した後から現在までの比較的温暖な気候で人類が急発展した時代の言語の多様性を理論的に探った研究で、7月25日 Science に掲載された。タイトルは「The rise and fall of language diversity through the Holocene(完新世での言語多様性の盛衰)」だ。
人間が生み出した道具と比べると、言語は音以外の物理性がない点で際立っている。この物理的制限がないおかげで、未来や目的など存在しないものを表現できる唯一の道具として、宗教から科学まで人間のあらゆる文化の基盤となってきた。ところが、この物理性の欠如は、文字が生まれる前の言語の研究を困難にしてきた。
この研究も含めて完新世の言語多様性の研究はほとんど理論的な研究になる。これまで3種類の仮説があり簡単に紹介しておくと、
Megadiverse paleolithic hypothesis : 農耕により集団が大きくなる前に、人間の集団数だけ言語が生まれ、その数は何万にも達したが、農耕が始まってからはコンスタントに減ってきた。
Demographic Boost Hypothesis : 逆に農耕は人口動態を安定化させ、これに呼応して言語数も増加、農耕前に数千に達していた言語が人口とともに1万程度に自然増加した。
Premodern equilibrium hypothesis : 言語の数は、植民地主義以前はずっとコンスタントに1万近くで維持されていた。即ち、農耕などはほとんど言語多様性に影響がなかった。
それぞれの仮説の基盤となるエビデンスがあるのか全く知らないが、大議論が続いていると言った話ではないように思える。これに対し、この研究では民俗学や人口動態研究に基づいて仮説を立て、言語多様性の変化を調べたのが、この研究の重要性だと主張している。
この研究では、言語は集団で生活する狩猟採取民から発生し、従って狩猟採取民の数が言語の数とほぼ同じであること、農耕による定着前の完新世での狩猟採取民のグループサイズは一定化していたこと、しかし、農耕などにより急速に人口が増加し、言語数の増加を遙かに上回るようになった。以上のことを基盤に、モデルを作成、言語数を推定している。
これによると、農耕前に5000近くに達していた言語多様性は、農耕による人口増加とともに、さらに拡大を続け、これまで言語数を低下させると考えられていた エジプト、メソポタミア、黄河 などの文明の誕生後もさらに多様性が加速し、2000年前にピークに達したと結論している。即ち、これらの文明は他の地域を一つの言語圏へ集約させるモーメントはなかったと考えている。しかし、2000年になると、これまでの仮説が全く及ばない要因が発生する。例えば世界宗教、ローマなどの植民地主義等などだ。そして、ヨーロッパ列強による植民地主義が広がる500年前ぐらいから、言語数は急速に低下し、現在は農耕前の7000程度の言語数に低下した。
以上が結果で、ある程度納得は出来るおもしろい研究だとは思うが、まだまだ本当かという疑問の心は残る。また、この研究の基本は完新世の初期段階が中心で、文字のインパクトはあまり考慮されていない。しかし、物理性のない言語に物理性を付与する文字は、文字のない言語が消失する大きな要因になったのではと想像する。
こうして考えると、そのオリジンはわからないが、最終的に1億人が日本語で統一され、しかも漢字を日本語用に使い回して日本語に同化させ、ワープロの登場で漢字を使うことが楽になった日本語の歴史は奇跡に感じられる。
2026年7月24日
感染性のガンの存在はイヌを飼っている人には広く知られていると思う。主として交尾中にイヌからイヌへ伝搬し、遺伝子解析からオオカミから存在し、数百年以上にわたって維持されていることが示されている。イヌ以外には、このブログで何度も紹介したタスマニアデビルの顔面に発生する腫瘍で、食べ物を争うときに感染し、一時はデビルは絶滅の危機にまで追い追い詰められた(https://aasj.jp/news/watch/4641 )。これに加えて北アメリカのオオノガイの白血病は、細胞が海を漂って他の貝にに感染することが報告されている(https://aasj.jp/news/watch/3246 )。
今日紹介する米国・バーモント大学からの論文は、バーモント州の Memphremagog 湖のナマズに発見されたメラノーマが感染性であることをゲノムから証明した研究で、4番目の感染性腫瘍の発見になる。タイトルは「Brown bullhead catfish melanoma represents a novel transmissible cancer ブラウンブルヘッドナマズのメラノーマは新しい伝搬性のガンだ」」で、7月19日 Nature にオンライン掲載された。
この湖では2012年に一部のブラウンヘッドナマズ (BHC) にメラノーマが見つかることが報告された。そして、2014年から2017年にかけて、なんとメラノーマにかかった個体が23-37%へと上昇し、一般的な個別の個体に発生するメラノーマの頻度をはるかに超えていることがわかった。そこで、第4の感染性腫瘍の発見と考え、19個体のメラノーマと正常脳組織、さらに近くの湖の BHC の全ゲノム解析を行っている。
まずミトコンドリアゲノムを調べると、全てのメラノーマはホスト組織と異なっていること、そして腫瘍のミトコンドリアは多様化はしていても、は同じ起原を持つミトコンドリアに由来していることが明らかになった。
次に核染色体のゲノムを調べると、メラノーマと、正常組織の系統樹は全く異なることが明らかになった。即ちメラノーマの起原は、この湖で見られる個体のゲノム多様性が生まれるより早い段階で発生し、その後メラノーマ自体で多様化していることがわかる。さらに、組織でもメラノーマでも一塩基レベルの多様化が発生しているが、個体で見られる多型のほとんどは1個体だけでしか見られないが、メラノーマの多型の多くは16/19メラノーマを筆頭に、複数のメラノーマで共有されていることを明らかにした。
さらにより大きな構造変化を見ると、メラノーマではほとんどの変異が12種類以上の個体で共有されているが、脳組織では頻度も低く、ほぼ全てが1個体だけで見られる。
以上が結果で、ゲノム解析だけが行われているが、間違いなくメラノーマが感染性に伝搬していることは間違いないと思う。タスマニアデビルと異なり、野生の魚と言っても飼育することが出来るだろうから、ゲノムとガンの個体を超えた伝搬について実験的な研究へと進むことを期待する。このナマズは生殖時に身体をすりあわせるので、おそらくこの時に感染したと著者らは想像しているが、感染の拡大から考えると、実験的な感染実験と、それに基づく個体間感染のメカニズムが明らかに出来るのではと期待する。
2026年7月23日
実は昨日起床して隣の部屋に行くとき失神し、肋骨を骨折してしまいました。最初脱水による立ちくらみかと思っていたのですが、病院で血圧が低く、130を超す洞性頻脈があることを指摘され、心房細動による血圧低下が原因ではと、一晩病院に泊まり様子を見ました。たまたま炎症再生委学会で講演をする予定だったので、会長の椛島先生にお願いして病院からzoomで講演をしたのですが、初めて10分ぐらいで頻脈が見事に治ってしまいました。結局ホルター心電図を装着して退院する前にこの記事を書いています。肋骨骨折はCTでははっきりわからないのですが3本ほどカクカク言うので、間違いはないと思います。こちらの痛みの方がやっかいそうです。とは言え、今日もおもしろい論文を紹介することが出来ました。
真核生物と原核生物を比べた時、イントロンの存在は真核生物の特徴の一つだ。そして、遺伝子がイントロンを含む場合、必ずスプライシングによって翻訳される遺伝子を作る必要がある。このようなイントロンやスプライシングの進化を考える時、イントロンのない原核生物を形成して、イントロンの意義を調べるのは一つの方法といえる。
今日紹介する中国科学アカデミー上海生化学細胞生物学研究所からの論文は、イントロンは真核生物にとって必要かという素朴な疑問を、全てのイントロンを出芽酵母ゲノムから取り除くのに成功し、イントロンなしでも真核生物は生きていることを証明した論文で、7月23日 Cell にオンライン掲載された。タイトルは「A spliceosome-independent eukaryote generated by complete intron removal(スプライソゾームの必要ない真核生物をイントロンを完全に除くことで作ることに成功した)だ。
出芽酵母には300のイントロンが存在し、これらを一つ一つ欠損させるとすると、10年以上の月日がかかるらしい。そこで、ランダムにイントロンを導入した後、二つの染色体を持った酵母内での減数分裂時の組み替えでイントロンの除去数を増やす方法で、最終的に完全にイントロンが存在しない SYNE27 の作成に成功する。それまでの中間段階の解析もおもしろいが、全て割愛して SYNE27 について説明する。
スプライシングが必要無くなってさぞかし楽になったかと思いきや、イントロンが減るにつれて増殖力は低下していく。しかし、SYNE27 を正常のゲノムとディプロイドにしても細胞増殖は低下しないことから、増殖にネガティブに働く要因が存在するわけではない。
増殖が低下する原因を調べると、リボゾームの合成に関わる様々なリボゾームタンパク質の合成が低下し、翻訳活性が低下することが最大の要因であることがわかった。このことは、イントロンの獲得とスプライシングシステムが進化的に染色体を持つ真核生物の適応性を高めることを意味する。その意味で、イントロンの全く存在しない SYNE27 をスタートラインにして、イントロンの新たな獲得の意義を調べる新しい研究が可能になる。
イントロンを全部は除去しても、スプライシングに必要な遺伝子は全て残っている。しかし、SYNE27 からスプライシングに関わる様々な分子を除去しても、SYNE27 は増殖を続ける。即ち、スプライシング分子は、スプライシング以外の他の機能を保っているかもしれないが必須ではない。おもしろいのは、イントロンが存在しない SYNE27 でも、配列の中からスプライスシグナルを探し出して遺伝子の一部を除去する反応が低い確率であるが見られる。これはイントロンのある酵母では見られない反応なので、本来のスプライシング標的がなくなったときに現れるが、原始的なスプライシング酵素がどのような mRNA 調節を担っていたのかを知るための重要なヒントになるかもしれない。
以上が主な結果で、最後に書いてあるが4年という月日を費やしてイントロンが全く存在しない出芽酵母を作成したことは、本当に素晴らしいチャレンジだと思う。さらにこの過程で、様々なレベルでイントロンが除去されて中間段階の酵母が樹立され、また SYNE27 ではスプライシングに必要な分子を全て除去する株の確率にも成功している。これらの材料は、スプライシングの進化を考える上で必須のツールになる。これまで、遺伝子の中に飛び込んだ selfish DNA がイントロンの起原と考えられているが、この研究で 自己的 DNA がホストに取り込まれて、安定な遺伝子制御に使われた可能性が示唆されている。一つの合成生物学の道筋がついた気がする。
2026年7月22日
組織標本上で何千ものRNAやタンパク質の発現を調べたり、あるいはトランスクリプトームを調べたりできる、空間トランスクリプトーム法についてはこのブログでも何度も紹介し、おそらく実験室でも高額ではあっても利用可能なテクノロジーになってきている。そして、次から次へとより困難な課題にチャレンジする研究も生まれている。
今日紹介するシカゴ大学からの論文もこのようなチャレンジの一つで、複数の抗体による染色とVisiumトランスクリプトームを組み合わせて、分子間、あるいは細胞間相互作用が起こっている場所でのトランスクリプトームの変化の解析を可能にする研究で、7月17日 Cell にオンライン掲載された。タイトルは「Spatial proximity sequencing maps developmental dynamics in the germinal center(トランスクリプトームの空間的近接性は胚中心の発生動態を明らかにする)」だ。
胚中心はリンパ組織内で免疫反応に関わる構造で、抗原刺激により現れ、抗体産生とその成熟に必要なリンパ球や樹状細胞の相互作用が進むダイナミックな組織だ。この研究の目的は、複数の抗体を用いて分子間あるいは細胞間の相互作用を特定した後、そこで起こる遺伝子発現変化を解析する方法の開発なので、胚中心は方法を検証するのに格好の対象と言える。
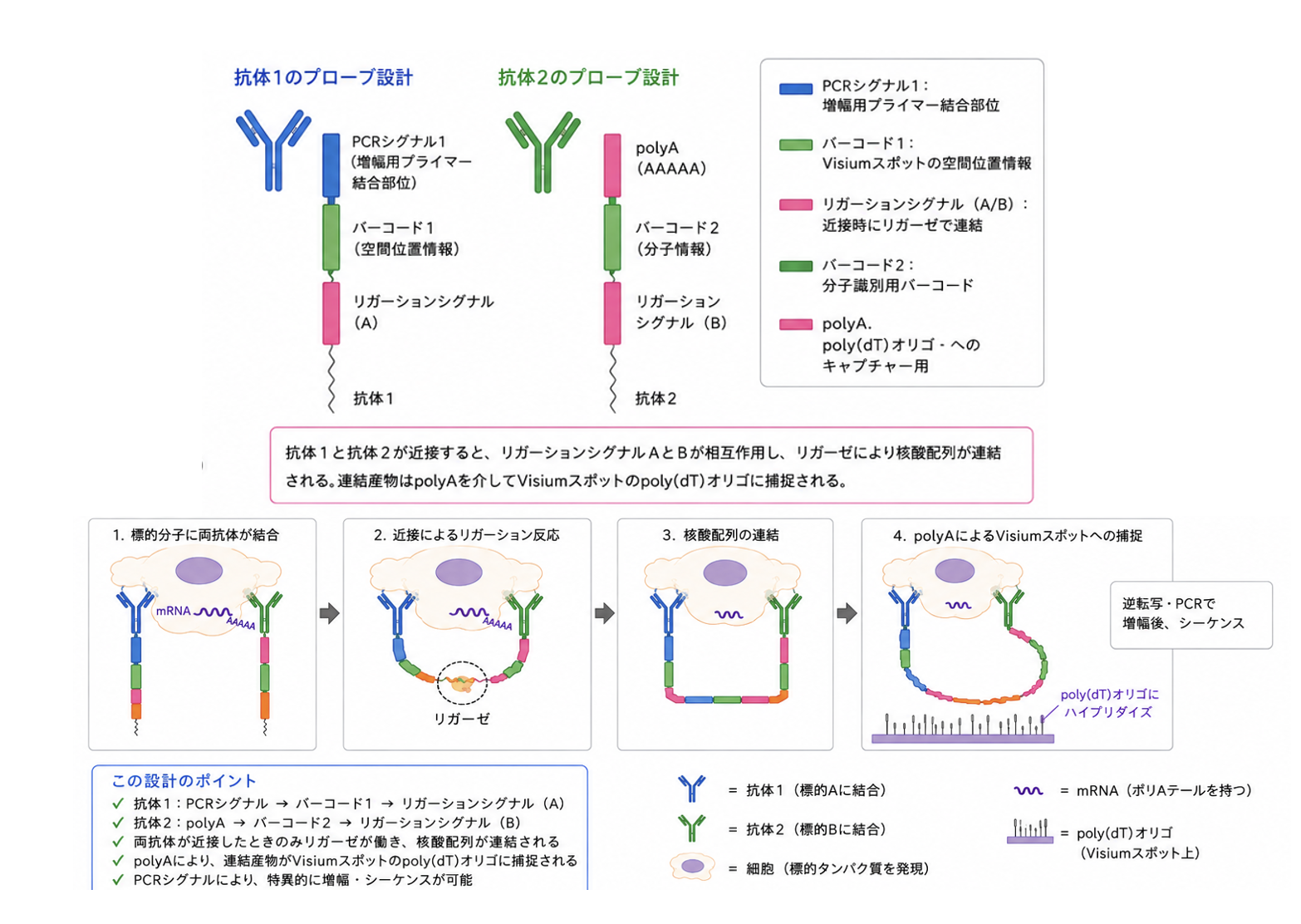
さて方法について詳しめに解説する。この方法の基盤は 10XGenomics 社の Visium と呼ばれる空間トランスクリプトーム法で、2ミクロン平方のスポットに異なるバーコードを持つ polyT を結合させ、その上に貼り付けた細胞に発現する mRNA をキャプチャーし、そのあとスライドグラス上の全ての RNA をまとめてシークエンスしたあと、バーコードに従って発現していた RNA の空間配置を再構成する方法だ。
この研究で開発された Sprox-seq と呼ぶ方法は、これに32種類の抗体を用いる染色を合体させる方法だ。重要なのは抗体を組み合わせるだけでなく、分子と分子が近接して相互作用している現場を特定することを目的としている点だ。事実、抗体を Visium に組み合わせるだけなら、抗体に PolyA と特定する配列を結合させておけば、発現 RNA とともにどの抗体が結合していたかを、同じ配列解析から抽出することができる。一般に行われるかどうかはわからないが、Visium 自体は RNA だけでなく、原理的には抗体によるタンパク質の同定も同時に出来る立て付けになっている。
例えば接着分子同士の結合を考えると、2種類の抗体が同じスポットに現れれば可能性は高いと推察できるが、Visium の解像度では本当に近接していたのかはわからない。そこでこの研究では、Aと言う分子に対する抗体に一つの PCR シグナルとバーコード、そして DNA 同士のリゲーション部位を結合させ、Aと反応するB分子に同じくリゲーション部位、バーコード、そして Visium にキャプチャーされるための polyA を結合させ、両方の抗体が近接する場所だけ抗体の情報がスポットに残るようにしています(この文章を読ませて Chat に書かせた図を掲載しておきます)。
これによって、例えば接着分子が結合している部位を空間トランスクリプトーム上にマップすることができる。
様々なテクニカルな条件を検討した後、胚中心で相互作用している分子の局在や、結合の強さを調べているが詳細は省く。代わりに一つだけ例として CD29 と VCAM1 、インテグリンα4 の接着分子同士の結合を検討し、インテグリン α4/VCAM1 及び CD29/VCAM1 の結合の局在が異なることを示している。
方法を説明したかったので、胚中心のバイオロジーについてはほとんど割愛してしまったが、発生学、免疫学、腫瘍学など様々な分野で役立つのではないだろうか。しかし、空間トランスクリプトーム解析法の進展は著しい。
2026年7月21日
細胞レベルでも、臓器レベルでも古くなった分子や細胞を新しいものに置き換えることが生命にとって必須の過程だ。この過程の異常は老化の重要問題として研究されてきた。まずリクルートする側、即ち幹細胞から分化した細胞の供給が老化で低下することは、だれもが感じる。さらに、最近では老化した細胞を速やかに除去できないと、臓器や個体全体の老化が促進されることが示され、死にかけの細胞の処理を速めるセノリシスによって老化を食い止める方法の開発が進んでいる。ただ最近様々な治験結果が出てきて、人間でセノリシスが老化を抑えるという仮説については再検討が必要なようだ。
今日紹介するスタンフォード大学からの論文は、古くなった細胞を処理する機構を高めて老化を防ぐという点ではセノリシスのアイデアに似ているのだが、処理される細胞が好中球に限られており、古くなった好中球が臓器に溜まると障害を起こすが、これが臓器老化の一因であることを示す研究で、7月16日 Science に掲載された。タイトルは「Restored clearance of senescent neutrophils by tissue-resident macrophages limits organ aging(老化した好中球の除去機能を組織で回復させると臓器の老化を抑制できる)」だ。
この論文はおもしろいというより、老化という定義の難しい言葉を使ってしまうことの問題を認識させる論文だとおもい紹介することにした。肝臓のクーパー細胞、脳のグリア細胞など、組織内で再生して機能するマクロファージは、tissue resident macrophage (TRM) として知られている。
この研究に至ったいきさつはわからないが、まず TRM 特異的にプロスタグランディンE2受容体 (EP2) をノックアウトするところから研究が始まっている。TRM とそれ以外のマクロファージを区別することはそう簡単ではないので、完全に TRM 特異的 KO とは言えないが、TRM での発現レベルが半分以下になるマウスを作成して、肝臓、肺、脳で TRM を調べている。TRM は老化とともに数が低下するが、EP2-KO ではこの低下が抑制される。
特に自然炎症や免疫に関わる遺伝子発現を見ると、老化 TRM で上昇傾向が見られるが、これが EP2-KO で元に戻る。これは、肝臓や肺だけでなく、腸、心臓、腎臓でも見ることができる。大事なのは、その結果で老化に伴う脳の認知機能低下が防がれ、四肢筋肉が強くなり、心筋の強度も高まる。実際に TRM の数の低下は2-3割程度で、それが9割ほど元に戻るだけなのだが、臓器機能への効果は意外に高い。
この原因を single cell RNA sequencing で調べると、老化によるTRM の減少に呼応して、老化した好中球や、細胞死の途中の好中球が処理されずに溜まってきていることがわかる。それだけでなく、溜まった死にかけ、あるいは細胞死細胞から炎症性サイトカインが放出され、NETosis と呼ばれる核酸の排出まで起こっている。ところが、PE をノックアウトすると、これらの異常がほとんど正常化することがわかった。このように、死にかけの好中球を処理することをエフェロサイトーシスと呼ぶが、老化に伴い TRM の数が減るだけでなく、エフェロサイトーシス機能が低下し、その結果死にかけの好中球から炎症物質が出て臓器を傷害する。
老化に伴う TRM のエフェロサイトーシス機能低下は、細胞をマクロファージが取り込むときに必要なインテグリンなどの発現異常によっており、EP2-KO でこれら機能分子の転写調節に関わる分子の発現が活性化されることで、エフェロサイトーシスが正常化することも示している。またノックアウトだけではなく、EP2 機能を阻害する化合物を2ヶ月投与する実験で、肝臓や脾臓で老化とともに上昇してくる死にかけの好中球の数を、化合物により低下させられることも示している。
最後に、人間の肝臓の single cell RNA sequencing データベースから、人間でも老化に伴い死にかけの好中球が増加し、炎症性サイトカインの分泌も高まっており、マウスと同じ状態が起こっていることを示している。
結果は以上で、元々プロスタグ TRM を減らし、マクロファージのエフェロサイトーシス機能を落としてしまう結果、死にかけの好中球が臓器に溜まることで、逆に炎症が起こるという結論だ。そして、EP2 機能を抑えることで、老化を防げるという結論になっている
しかし、これを老化と言っていいのかわからない。EP2 も若い段階では炎症が高まらないよう重要な機能を果たしている。しかしこの論文では追求し切れていない、年齢による影響により TRM 特異的に思いがけないサイクルが始まり、この時 EP2 のような炎症を抑える分子が逆に炎症を高め、臓器が傷害されるという話だ。これを老化と一言でまとめていいのか、そして EP2 阻害を若返りの薬と言っていいのか、個人的には違っているような気がする。
2026年7月20日
ガンのゲノム解析の重要性は言うまでもないが、RNAの発現や免疫染色も重要だ。例えば私のガンの場合、発現が高い遺伝子としてアンドロゲン受容体とその下流分子が見つかるし、乳ガンで最も研究が進んでいる Her2 とそのファミリー分子も発現している。とすると、状況によっては変異はなくてもこれら分子を標的として使うことができる。
Her2 を標的に使う治療として最近大成功を収めているのが Her2 抗体にトポイソメラーゼ阻害剤や、微小関係性阻害剤を結合させた ADC 治療だ。もちろん抗体が結合する必要があるので、Her2 の発現が高い必要があるが、第一三共のエンハーツは Her2 発現が低い場合も一定の効果があることを証明し、この薬剤の宣伝文句になっている希望を生み出している。しかし、エンハーツでも Her2 の発現が高い方が効果は高い。
今日紹介するワシントン大学からの論文は、エンハーツや同じ Her2 抗体に微小管阻害剤を結合させたカドサイラを、高い確率で Her2 と同時に発現している EGFR を利用することでリソゾームへの取り込みを高めて、Her2 の低い場合でも効果を高める方法の開発で、7月15日 Nature にオンライン掲載された。タイトルは、「Modular in vivo antibody–ADC click to reverse drug resistance in tumours(抗体とADCを生体内で組み合わせるクリックを用いて腫瘍の薬剤耐性を抑える)」だ。
私の場合もそうだが、Her2 発現のガンは EGFR も発現していることが多い。エンハーツもカドサイラも Her2 に結合して細胞内に取り込まれて薬剤が外れるので、Her2 の発現が多い方が良いし、また細胞内に取り込まれることも重要になる。この時、EGFR が Her2 と結合すると、Her2 の細胞内への取り込みを抑えることがある。
そこでこの邪魔な EGFR を薬剤を取り込みヘルパーにしようと考えたのがこの研究だ。方法だが、EGFR に対する抗体と、ADC (エンハーツやカドサイラ) に、生体内でそれぞれの抗体を結合させることが出来るクリックと呼ばれている小分子を結合させる。まず、EGFR に対する抗体を投与した後、クリック結合可能な ADC を投与すると、EGFR と Her2 を発現するガン細胞上で複合体が形成され、Her2 だけでなく EGFR によっても細胞内へ取り込まれることが期待できる。
試験管内でこの可能性を確かめた後、クリックADC をアイソトープラベルしてマウスに投与すると、クリックで結合可能な EGFR を前もって投与していた群のみ、腫瘍内へのアイソトープの取り込みが高まる。これを Her2 の発現が高いガンで確認した後、今度はHer2の発現が極めて低いエンハーツだけでは効果が低いガンで調べると、ADC がガンに効率よく取り込まれ、単独治療よりはるかに強いガン抑制効果を得ることができる。
さらに、EGFR の発現が高まった結果 ADC が効かなくなったガンに対するクリックADC の効果を調べ、クリック・エンハーツがほぼ完全に腫瘍を抑制できることを示している。
最後により安定なクリック化合物の抗体への結合方法として、抗体や ADC の糖鎖にクリック化合物を投与する方法を開発し、これにより極めて均質にクリック化合物が結合した EGFR 抗体と、ADC の組み合わせを作れること、これを用いてエンハーツだけでは効果がなくなった膵臓ガンや乳ガン細胞の増殖を抑制できることを示している。
以上が結果で、Her2 が ultralow でも EGFR が発現しておればガンを強く抑制できると言う結果は、私たちの希望になることは間違いないが、しかし正常細胞にも Her2-lowEGFRhig は存在するので、ヒトでの治験が待たれる。さらに、この結果はこのようなクリックの利用による2種類の抗体を結合させることで、これまで難しかった多くのガンをたたける可能性を秘めている。期待したい。
2026年7月19日
タンパク質のデザインに AI が使われるのは珍しくもなんともない。しかし AI の専門家ではない生命科学の大御所が AI を利用した論文は、なかなか味がある。例えばエピジェネティックス研究の大家 Richar Young が ESM2 を利用してタンパク質の細胞局在を予測する方法の開発(ProtGPS: https://aasj.jp/news/watch/26318 )では、大御所ならではの明確な研究目的の設定など、生成 AI もプロが使うようになるとさらに大きな発展があることを予感させた。
今日紹介するのは遺伝子編集の大御所中の大御所 Doudna さんが、コンパクトな遺伝子編集分子を AI を用いてデザインした論文で、AI と専門知識の共同することの重要性を見事に示した研究で、7月16日 Science に掲載された。タイトルは「Structure and evolution-guided design of minimal RNA-guided nucleases(最小限のRNAガイドによるヌクレアーゼを構造と進化をベースにデザインする)」だ。
遺伝子編集には細胞への導入効率の点で、遺伝子をできるだけ小さくすることが重要になる。この点で、Doudna さんたちが開発してきた CRISPR/Cas システムより小型のシステムとして、Cas12 の祖先筋のトランスポゾン TnpB が注目されている。この研究の目的は、現存する TnpB を人工的にデザインしてより小さな RNA/DNA 配列を認識して遺伝子を切断する人工酵素を作ることだ。
これまでの生命科学では、これを実現するため既存の TnpB に変異や欠損をランダムに導入する人工進化システムが使われていたが、この役目を生成AIにやらせて見ようというのが今回の研究だ。現存の生命のゲノムに残っている進化経路以外にも、多くの潜在的進化経路が存在し、それを生成 AI が確率空間として表現しているとすると、Doudna さんが AI を利用しようとするのも当然の成り行きだ。
このために彼女が選んだ AI が ESM-IF と呼ばれる2022年に開発されたモデルで、タンパク質の構造を入力すると、アミノ酸配列が出力されるというアルファフォールドの逆モデルだ。おそらく、TnpB や Cas の構造を熟知した Doudna さんならではの選択に思える。
ただ使ってみると、Doudna さんの期待は裏切られ、彼女がこの分子の機能に最も重要と考えるアミノ酸まで自由に変えたアウトプットが出てくることに気づいた。そこで、構造だけでなく、実際に進化過程で保存されたアミノ酸部位を条件として与えて、この制限の範囲内で自由に設計できる方法を考えた。実際には、TnpB としての保存度 (Co) と、TnpB が様々な配列の RNA や DNA に合わせて共進化する coupling strength をσとして、この閾値を研究者が決めることで設計の自由度を制限する方法を開発した。この coupling strength 等はまさに遺伝子編集分子を熟知した Doudna さんならではの境地だと思う。
さらに、RNA/DNA 結合ドメインとヌクレアーゼドメインを一体としてデザインさせると、相互に作用し合って設計が難しくなることを考慮して、それぞれのドメインの設計を、実際の TnpB の持つもう一つのドメインと結合させて制限するインプットで行わせ、現存する TnpB と比べて72-83%という多様性を持つ人工 TnpB を生成させることに成功している。ここにも専門家の視点が生きている。
こうして生成した TnpB の機能はバクテリアの生存としてチェックできるので1万個を超えるアウトプットでも評価が可能で、最終的に実際の TnpB と同じかそれ以上の編集効率を持つ9種類の人工 TnpB の設計に成功している。
この人工 TnpB を用いてヒト腎臓細胞株での遺伝子編集効率を、実際の TnpB と比べ、条件によっては5倍以上の効率で働くことを明らかにしている。その上で、何故人工 TnpB で効率が高いかをクライオ電顕で構造比較し、新しくデザインされた分子では、より核酸との結合が安定化するアミノ酸が導入されていること、さらに TnpB の進化過程では捨ててしまった TAM-結合部位が復活していることを発見する。まさに、進化可能性の潜在空間から新しい分子を生成出来ることを見事に示した。
結果は以上で、Doudna さんの脳という遺伝子編集知識の集大成が、ESM-IF の足りない点を補うことで、それぞれでは難しい課題が解決されたことが示された素晴らしい研究だ思う。このようにトップ研究者ほど AI を生かし、新しいモデルも開発できることを考えると、ただ闇雲に AI 研究者を生命科学に動員すればいいという話ではないことがわかる。まさに、それぞれの研究現場で利用のためのチャレンジを進めることが重要なのだ。
とは言え、Doudna さんの頭の中にしまわれている遺伝子編集分子の構造や進化の知識を、将来 AI に置き換えることも可能だろう。例えば原核生物の進化潜在空間を学習した Evo1 などを ESM-IF と組み合わせることで、専門家以外にも課題解決が可能になると確信する。しかしそのためにも、AI と専門家の共同が重要だと認識した。
2026年7月18日
利根川先生の訃報を聞いたので、新しく Science に掲載されていたカロリンスカ研究所から発表されたγδT細胞を刺激する自己分子の話を取り上げることにした。
大学卒業後、免疫学から始めたので、利根川先生についての思い出は多い。最初はまだ研修医の頃、京都大学胸部疾患研究所の会議室で、凱旋帰国した利根川先生の免疫グロブリン再構成の可能性についてセミナーが行われ、同級生の武藤君と聞きに行った。まだ southern blotting と言う時代ですらなく、DNAの会合・再会合を調べる Cot 解析で、リンパ球のDNAが生殖細胞と異なることを示した話だった。
その後1980年に留学した時は、ちょうど MIT に移られる頃だったが、バーゼル研の利根川研で夜昼なく働いていた同世代の黒沢さん、坂野さんと知り合いになり、帰国後も付き合いは続いた。バーゼル研での利根川さんの様々な逸話は彼らから聞かされたので、いつか彼らの口から語られるのを待とう。
免疫グロブリンの研究は1970年代の話だが、1980年当時はT細胞受容体遺伝子を明らかにすることだった。T細胞受容体遺伝子をだれが最初に明らかにするのか、当然利根川先生が本命と考えられていた。ところが京都で行われた免疫学会が最初だったと思うが、全くダークホースと言える Mark Davis や Tak Mak が T細胞受容体遺伝子のクローニングに成功した。
この競争の中でもう一つ明らかになったのが、全く新しいγδT細胞で、通常の αβTCR遺伝子とは異なるγδTCR遺伝子を再構成して表現するおもしろい T細胞で、最初に利根川先生により報告された。そんな訳で、γδT細胞とノーベル賞のカロリンスカ研究所が利根川先生と結びつき、この論文を紹介することになった。
利根川先生の発見以来40年以上経つが、この細胞についての謎は多い。中でも、いくつかのγδT細胞が胸腺で自己分子に選択され、Th1T細胞へ分化するという発見だ。とはいえ、ほとんどのγδV遺伝子組み合わせに対応する自己分子はほとんどわかっていない。その中で、なんと炎症に関わる IL-17受容体を新しいγδT細胞刺激自己分子として発見したのがこの研究で、7月16日 Science に掲載された。タイトルは「Direct interaction of Vδ7 TCRs with IL17RA drives the differentiation of TH 1-like γδT cells(Vδ7TCRとIL17RAの直接相互作用がTH1様γδT細胞の分化を誘導する)」だ。
この研究では胸腺から樹立した細胞に反応する TCR を探索し、調べたほとんどの TCR を刺激する胸腺リンパ腫細胞を発見する。即ち、自己分子がいくつかのセットのγδTCRと反応する。次に反応している分子を特定するため、この細胞の遺伝子をクローニング、その中から TCR を刺激する分子を探索して、IL17RA分子を特定する。
次に、IL17RA分子をラベルして、これに結合する T細胞を集め、その TCR遺伝子を調べると、全て例外なく Vδ7 を発現しており、γの方は Vγ4 か Vγ1 である事がわかった。おもしろいのは、Vδも再構成部位には多くの変異が入っており、これが IL17RA との結合多様性に関わる点だ。おそらく、CD3領域の変異は分化後に導入されていると思うので、今後この変異の起こり方の理解が重要な課題になると思う。
ただこの研究では機能より、IL17RA と Vδ7 の結合に関わる部位、及びこのT細胞の発生に焦点を当てて研究している。構造的には CDR2 を挟む FR2、FR3 領域が直接 IL17RA に結合する。即ち、例えば HLA のような抗原を提示する分子は全く必要がなく、直接結合する。
多くのγδT細胞は腸管に分布するが、このタイプはほとんど脾臓やリンパ節に存在し、ほとんどが TH1 型の遺伝子発現を示す。そこで、γ4δ7TCR を持つトランスジェニックマウスを作成し、IL17RA抗原を発現するマウスと掛け合わせると、このタイプの T細胞が胸腺の CD4-CD8-細胞段階から発現し、ここで IL-17RA と反応して TH1 への分化のシグナルが入り、末梢へと移動することを明らかにしている。この過程は、IL-17RA のノックアウトされたマウスでは起こらず、またγ4δ7T細胞の増殖も起こらない。即ち胸腺のポジティブセレクションを IL17RA が誘導し、そのまま TH1細胞へと分化させている。
結果は以上で、自然炎症に重要な IL-17RA がγδT細胞を誘導する自己分子として働いているという驚くべき結果だが、何故このようなγδT細胞系が存在するのか、あるいは人間にも存在するのか、そしてその機能は何か、等まだまだ謎は続く、
2026年7月17日
今日はどちらも Nature Medicine に掲載されていたアルツハイマー病 (AD) についての臨床研究を紹介する。
最初のワシントン大学からの論文は 環状 RNA を用いて AD のステージについての正確な診断を行う試みで、7月1日にオンライン掲載された。タイトルは「Blood-based circular RNAs for early diagnosis of Alzheimer’s disease(血中の 環状 RNA はアルツハイマー病の初期診断に使える)」だ。
これまで症状の出る前から AD のリスクを判定できる血中マーカーとして pTau217 が開発され、広く使われるようになった。ただ、アミロイド沈着に依存するため、抗体治療でアミロイドが除去されると病気のモニターに使えなくなる。この研究では神経細胞由来の RNA 、とりわけ血中で安定な 環状 RNA を用いてAβや Tau の治療にかかわらず診断が出来ないか、500人近くの AD のコホート研究サンプルを用いてチャレンジしている。
結果、主に脳で発現している mRNA がスプライシングを受ける際に発生する34種類の 環状 RNA の量を組み合わせた指標を開発し、この指標が アミロイドPET 等の他の検査と比べても、AD リスクや病態と相関する検査になることを示している。pTau217 と組み合わせて使うと、AUC で0.931と言う高い特異性で アミロイドPET 陽性の患者さんを発見できることを示している。最初、神経が損傷されて 環状 RNA が出てくるから神経変性疾患の間で特異性がないのではと思ったが、なんと AD 特異的で、逆に言うと他の神経変性疾患でも同じようなマーカーを開発できる可能性がある。
環状RNA は認知症が始まる2年前から陽性になり、この数値は予後と強く相関する。更には胃や胃段階から上昇する pTau217 と比べるとその精度は一段と上がる。
以上が結果で、ここでは検討されていないが、アミロイド抗体を用いた治療群を調べると、検査としての重要性がより明確に示されると思う。実際にはシークエンスベースで行っているが、簡易検査として確立することを期待している。
次は米国の Immunobrain Checkpoint Inc という会社と、イスラエルワイスマン研究所などからの論文は、軽度認知を示す AD 患者さんの進行を、ガンのチェックポイント治療に用いられる PD-L1 抗体を用いて抑える第一相の治験で、7月15日にオンライン掲載されている。タイトルは「Immunotherapy with a short-lived anti-PD-L1 antibody in Alzheimer’s disease: a phase 1b, randomized, double-blind trial(半減期の短い anti-PD-L1 を用いたアルツハイマー病の免疫治療:無作為化、2重盲検第一相b治験)」だ。
AD にチェックポイント治療を行う理由は、高齢のAD患者さんでは免疫機能が低下しており、ミクログリアなどの活性が低下して異常が拡大する傾向にあると考えているからだ。これは年齢とともに自然炎症が亢進することが AD のリスクになるとする考えの真逆になる。ただ、チェックポイントは獲得免疫の話なので、自然炎症とは別に獲得免疫が AD の進行を抑えているという前臨床研究が行われてきたのだと思うが、全く把握しておらず、こんな治験が始まっているのかと正直驚いた。
とは言え、ガンに使う抗体とは異なり、わざわざ半減期が短い抗体を設計し、しかも12週に1回と、かなりの間隔を空けて注射している。ただ投与量は30mg/kg と多く、例えば PD1 抗体は3mg/kgで使うことを思うと、短い期間免疫疲弊を戻すという方法だ。
結果だが、量は多くとも、間隔が短く半減期も短いので、通常のチェックポイントよりはるかに副作用は少なく、一番重い副作用として自覚症状のない肝機能低下が挙げられている。また、薬剤動態だが4日で抗体価が半減している。
効果だが、活性化T細胞が投与後1週間をピークに末梢血に出現し、免疫反応では期待の効果を上げている。そして、AD のバイオマーカーとしての脊髄液中の neurogranin、tTau。pTau の量は低下している。症状については、ほぼ一年の経過観察でホトンの大きな変化がないという結果だ。
なんとなく効いてそうな感じだが、軽度認知症を対象にAβ抗体を選ぶのか、PD-L1を選ぶのか、もっと大規模な第2相、第3相の結果待ちになる。いずれにせよ、ADも搦め手からの治療法はまだまだありそうだ。